УМОЗРИТЕЛЬНЫЙ АНАЛИЗ В ЭВОЛЮЦИОННОЙ НАУКЕ
|
|
|
|
Обзор литературы |
|
|
Современное состояние эволюционной науки |
|
|
Современные методы эволюционной систематики |
|
|
Обзор основных источников |
|
|
Выводы |
|
|
Материалы и методы |
|
|
Материалы |
|
|
Умозрительный метод |
|
|
Выводы |
|
|
Результаты и их обсуждения |
|
|
Принципы умозрительного метода |
|
|
Классификация ПУМ |
|
|
Принципы действия метода |
|
|
Принципы определения |
|
|
Принципы систематизации |
|
|
Выводы по параграфу |
|
|
Умозрительный анализ теории эволюции: Уровни и механизмы эволюции |
|
|
Уровни эволюции |
|
|
Особенности микроэволюции |
|
|
Понятие и механизмы мезоэволюции |
|
|
Понятие и механизмы мегаэволюции |
|
|
Особенности сингенеза |
|
|
Эволюция процесса размножения |
|
|
Взаимовлияние эволюционных механизмов |
|
|
Критика концепции искусственного отбора |
|
|
Выводы по параграфу |
|
|
Умозрительный анализ эволюционной систематики: Возможные пути эволюции жизни |
|
|
Происхождение прогенота |
|
|
к разделу 3. 3. 1. |
|
|
Происхождение и эволюция вирусов |
|
|
Происхождение и ранняя эволюция эукариот |
|
|
Эволюция Tubulicristata |
|
|
Эволюция Lamellicristata |
|
|
Горизонтальная эволюция беспозвоночных |
|
|
Вертикальная эволюция беспозвоночных |
|
|
Эволюция Radialia |
|
|
Выводы по параграфу |
|
|
Выводы |
|
|
Общие выводы |
|
|
|
|
|
|
|
|
Диагностика царств живых организмов |
|
|
Перечень высших таксонов современных организмов |
|
|
|
Современная наука переживает период тотальной специализации и математизации. С одной стороны, наблюдается наплыв методов из точного крыла фундаментальных наук в гуманитарное через естественнонаучный центр: практически поглотив такие в древние времена естественные науки, как геометрия и физика, наполнив химию, геологию, астрономию и описательные отрасли биологии, математика начинает уже проникать в гуманитарные науки. Находясь на пересечение фундаментальных биологических дисциплин, эволюционная систематика и теория эволюции также подверглись внедрению математических методов, что повлекло, однако, к значительному уходу от первоначальных целей биологии. С другой стороны, в самой биологии наблюдается тенденция к развитию вглубь, то есть ко все более детальной специализации на узких проблемах и отраслях, далеких от познания живой природы как уникального феномена. В эволюционной систематике это проявилось в увеличении количества исследований мелких таксонов при практически полном пренебрежении филогенией крупных: выявив родственные связи между разными видами отдельных родов, ученые до сих пор не могут прийти к общему мнению о количестве царств живой природы, их составе, эволюционных отношениях и критериях выделения.
Многие моменты эволюционной истории живых организмов до настоящего момента являются “белыми пятнами” эволюции. Среди важнейших из них и такие значительные вопросы, как происхождение жизни вообще и на Земле в частности, вирусов, прокариотического и эукариотического типа организации генома, различных царств живых организмов, высших растений, многоклеточных животных, отдельных групп животных и растений, человека. Разумеется, только фактического материала (хотя и его порой не хватает) здесь не достаточно: необходим и его подробный анализ.
В настоящее время появилась острая необходимость в обобщении накопленного теоретического и фактического материала, однако если различного рода практические исследования доступны почти каждому человеку, имеющему соответствующую подготовку, то теоретизировать по общенаучным вообще и общебиологическим в частности темам способны достаточно редкие индивидуумы, обладающие соответствующим складом ума и высоким коэффициентом интеллектуальности.
В данной работе представлена попытка оформления умозрительных методов (практически потерявших свою роль в естественных науках со времен натурфилософии, но, тем не менее, актуальных и по сей день), их применения к эволюционной систематике и теории эволюции как наиболее важным дисциплинам теоретической биологии, а также умозрительных построений, касающихся возможных механизмов и путей эволюции. В целом работа носит не столько общебиологический, сколько натурфилософский характер, хотя основывается в значительной степени на биологическим материале и предназначена для реализации биологических задач.
Целями данной работы является доказательство актуальности и жизнеспособности умозрительных методов в эволюционной систематике и теории эволюции и выявление с помощью умозрительного (дедуктивно-индуктивного) анализа основных механизмов эволюции и филогенетических отношений между крупными систематическими группами живых организмов. Для осуществления этих целей поставлены следующие задачи:
1. Краткое описание современного состояния эволюционной теории.
2. Определение, формулировка и систематизация основных принципов метода.
3. Выявление возможных механизмов эволюции и разделение макроэволюционного процесса на мезо- и мегаэволюцию.
4. Формулировка наиболее точного (полного и достаточного) определения жизни.
5. Создание наиболее достоверной гипотезы возникновения жизни на Земле.
6. Отслеживание основных этапов эволюции живой природы.
7. Составление возможной филогенетической системы живых организмов на уровне высших таксонов.
Данная работа, хотя и основывается на наиболее древних, умозрительных методах научного познания, может рассматриваться как новый подход к общетеоретическим проблемам биологии, поэтому даже в том случае, если отдельные выводы, сделанные в ходе этой работы, будут опровергнуты, умелое применение умозрительного анализа может дать огромный результат для развития фундаментальной науки в целом.
Многие факты, затронутые в работе, могут оказаться общеизвестными, однако их систематизация и сведение в единую работу сами по себе должны представлять интерес.
Мы выражаем благодарность Беляковой Надежде Вадимовне (Калининградский областной эколого-биологический центр учащихся) и Никитиной Светлане Михайловне (Калининградский государственный университет), без которых было бы невозможно возникновение наших нынешних эволюционных и общебиологических взглядов, Саблиной Нине Николаевне (МОУ - средняя общеобразовательная школа №10) - за предоставленную литературу, а также Владимиру Курову, Дмитрию Костикову, сотрудникам компьютерного учебно-методического центра МОУ - СОШ №10 и другим, оказавшим посильную техническую
поддержку при оформлении работы.§ 1. 1. СОВРЕМЕННОЕ СОСТОЯНИЕ ЭВОЛЮЦИОННОЙ НАУКИ.
В настоящее время биология переживает очередной кризис. Современный этап развития биологии характеризуется следующими отличительными особенностями. Во-первых, научно-технический прогресс привел к резкому возрастанию количества фактического материала практически по всем отраслям биологии, что не только позволило изучить ряд биологических структур, процессов и механизмов, но и повлекло за собой возникновение множества новых наук, в основном, занимающихся ультраструктурными, биохимическими и другими основами жизни. Во-вторых, биология вслед за другими естественными науками начинает подвергаться активной математизации, то есть возрастанию роли математических методов, и даже зачастую - к приоритету их перед традиционными. В-третьих, имеет место все большее углубление в узкоспециальные отрасли вплоть до экологических, физиологических или генетических особенностей какого-либо единственного семейства, рода, а то и вида, при весьма незначительной доле исследований по глобальным общебиологическим вопросам (наподобие глобальных концепций Вернадского, Любищева, Берга), что хотя и ведет к более глубокому познанию отдельных живых объектов, но отнюдь не способствует развитию биологических знаний в целом. Все эти черты имеют как положительные, так и отрицательные стороны, однако очевидно, что они могут быть благоприятны только для отдельных отраслей, а не для общебиологической теоретики.
Одним из наиболее важных направлений фундаментальной биологии является, безусловно, эволюционная теория. Более того, выяснение филогенетических связей и эволюционных механизмов должно быть основной задачей фундаментальной биологии, в противовес практической биологии, преследующей строго утилитарные цели наподобие решения экологических, медицинских и продовольственных проблем (в таком случае многие современные направления в биологии оказываются совершенно бесполезными без конкретного применения на практике либо увязки с эволюционными представлениями). Однако, несмотря на резкий рост фактического материала, эволюционная теория весьма недалеко продвинулась со времен Ламарка и Дарвина и страдает неполнотой и постоянными разногласиями, в том числе по такому важному вопросу, как единство или самостоятельность микро- и макроэволюции [Назаров, 91, стр. 3-10]. Наиболее интересными в трактовке надвидовой эволюции можно назвать, пожалуй, концепции ортогенетического толка, в разработке которых принимали участие такие ученые, как Л. Берг, Н. Вавилов, А. Любищев, С. Мейен [Московский, 99], но они, обнаруживая ряд важных закономерностей в эволюционном процессе, придают мало внимания естественным механизмам эволюции. При всем этом, как ни парадоксально, противоборство различных эволюционных концепций, даже умозрительных, весьма напоминает противоборство в религиозном мире, когда, вместо того, чтобы искать что-то общее и продвигаться по пути синтеза, адепты каждого течения претендуют на абсолютную правоту.
К сожалению, дать полный обзор эволюционных теорий в работе, не касающейся напрямую их сравнением, нельзя, поэтому интересующимся можно порекомендовать работу Назарова [Назаров, 91]. Хочется надеяться, сто со временем будет разработана общая теория эволюции [Назаров, 91, стр. 11-12], в которой будут учтены все имеющиеся разработки на этот счет.
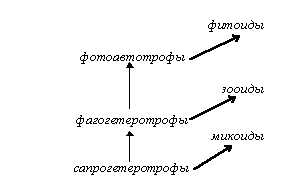
Рис. 1. Пример экологической системы (эволюция эукариот).
Эволюционная систематика также незначительно изменилась с начала века: современные биохимические данные имеются только по небольшому числу таксонов, они весьма мозаичны и не дают общей картины, - причем нет общего мнения даже по таким глобальным вопросам систематики и филогении, как происхождение жизни или эукариотического типа клеток, а также по количеству, структуре и эволюционным связям царств живой природы, выделяемых от двух (Прокариоты/Эукариоты или даже устаревшее Животные/Растения с абсолютно не обоснованным включением в последнее грибов, бактерий и даже вирусов [Хржановский, Пономаренко, 88, стр. 147]) до двух десятков и более [Маргелис, 83; Мирабдуллаев]. Интересно при этом, что, называясь филогенетической, современная систематика в значительной степени сочетает признаки экологической систематики (на экологическом принципе, в основном, построено разделение высших эукариот в традиционных системах на животных, растения и грибы, поскольку это деление, по сути, отражает не филогенетическое положение этих групп, а характер питания большинства их представителей: фотоавтотрофный у “растений”, или, точнее, фитоидов, сапрогетеротрофный у “грибов”, или микоидов, и фагогетеротрофный у “животных”, или зооидов (см. рис. 1), тогда как низшие эукариоты “скидываются” в царство Protista, или Protoctista [Маргелис, 83; Мирабдуллаев, 1]), систематики уровня организации (деление на прокариот и эукариот, высшие и низшие растения, простейших и многоклеточных, беспозвоночных и позвоночных, классификация позвоночных (см. рис. 2)) и систематики, основанной на морфологических особенностях (разделение “простейших” на жгутиконосцев и саркодовых, аннелид - на сидячих и бродячих, инфузорий - на ресничных и сосущих). На этом фоне серьезного внимания заслуживает концепция Любищева о систематике как “окне в онтологию”, согласно которой система живых организмов является не чем-то условным, а изначально “запрограммированной”, заложенной в природу системой, наподобие системы элементарных частиц или химических элементов [Московский, 99].
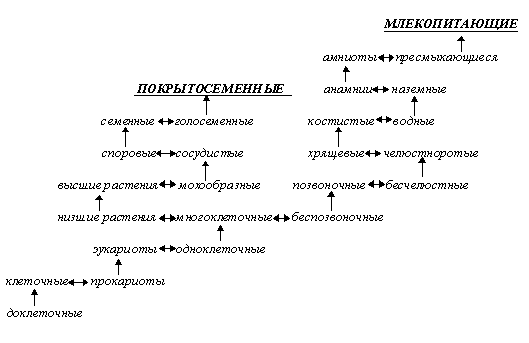
Рис. 2. Пример системы уровней организации (в рядах, ведущих к образованию покрытосеменных и млекопитающих).
Серьезную путаницу вносит и несогласованность между зоологической и ботанической систематикой и таксономией. Так, одни и те же систематические ранги могут называться по-разному в зависимости от того, к животным или растениям причислена та или иная группа (отряд или порядок, отдел или тип); латинское название одной и той же группы в зоологии и ботанике могут быть различны; могут меняться и систематические ранги той или иной группы в зоологии и ботанике (например, класс Растительные жгутиконосцы в зоологии соответствует целой группе мало связанных между собой филогенетически отделов/типов растений: эвгленовые, зеленые, желто-зеленые и другие водоросли).
Ликвидация, по крайней мере, явных нарушений эволюционного принципа в классификации живых организмов, таким образом, является одной из первостепенных задач современной систематики.
§ 1. 2. СОВРЕМЕННЫЕ МЕТОДЫ ЭВОЛЮЦИОННОЙ СИСТЕМАТИКИ.
Традиционные методы эволюционной систематики либо исчерпали свои возможности, либо оказались применимыми только в отдельных случаях. Сравнительная эмбриология, изучившая сходства в развитии представителей разных групп живых организмов, доказала также и то, что онтогенез - повторение не филогенеза, а лишь процесса эмбрионализации в филогенезе, да и то весьма упрощенное [Богомолов, 68; Грант, 80; Захваткин, 49; Зусман, 73; Рэфф, Кофмен, 86; Шпет, 77]; к тому же, она применима только в “средних пластах эволюции” (и то не всегда), то есть не раскрывает ничего или почти ничего, с одной стороны, о происхождении жизни, становлении прокариот и эукариот и формировании царств и, зачастую, типов, с другой - об эволюции внутри отрядов, а зачастую и более высоких таксонов, и сочетается с явлением филогенетического ускорения, когда, напротив, онтогенез повторяется в филогенезе. Интересным, но отнюдь не характерным исключением в этом плане является гипотеза колониального возникновения эукариот, согласно которой митотическое деление рассматривается как рекапитуляция процесса возникновения эукариот из колоний прокариотических организмов одного вида (противовес двум более известным гипотезам: симбиогенеза и мембранизации). Сравнительная анатомия, даже вкупе со сравнительной эмбриологией, физиологией, генетикой и эволюционными направлениями других дисциплин, явилась лишь доказательством самого факта существования эволюции, но не раскрыла ее механизмы и также оказалась беспомощна за пределами “среднего сектора”. Палеонтология, призванная, казалось бы, для подтверждения эволюции, в действительности практически полностью опрокинула дарвинистские представления о постепенном преобразовании видов, обнаружив скачкообразность их возникновения (сальтационное видообразование), на чем креационисты смогли строить достаточно убедительные доказательства Творения [Жизнь - как она возникла? 85, стр. 55-57], а биологи вынуждены были формулировать теорию квантовой эволюции, которая подтверждается только фактом ОТСУТСТВИЯ в палеонтологической летописи переходных форм, а не НАЛИЧИЕМ чего-либо, четко свидетельствующего о данном механизме эволюции (то есть, палеонтологическая летопись хотя и достаточно достоверно опровергает дарвинистские положения, но не доказывает ничего иного). Кроме того, вследствие понятной мозаичности летописи (даже то множество находок не может дать полной картины, тем более, о ранних периодах эволюции) данные палеонтологии могут носить характер только косвенных доказательств чего-либо. Биохимические, генетические и другие современные методы эволюционной систематики хотя и оказались весьма продуктивными, позволив выявить филогенетические связи между многими группами, ранее разнесенными даже по разным царствам, и, напротив, разделить ранее объединяемые виды, во-первых, не позволяют охватить все многообразие живой природы, во-вторых, дают, в основном, представления о степени родства, а не о направлениях и механизмах развития, и, в-третьих, все равно бесполезны без опытного теоретика [Маргелис, 83; Мирабдуллаев; Пахомова, 72]. Множеству же рассеянных по этим и другим наукам фактов, которые могли бы пролить свет на сущность и пути эволюции, просто не придавали еще эволюционного значения, и они пропадают без внимания.
С серьезными проблемами сталкивается и концепция тихогенетической (основанной на случайных изменениях) эволюции. В статье А. Московского “Существует ли научная альтернатива дарвиновской концепции эволюции?” в качестве основных проблем дарвинизма названы туманность формулировок, многообразие взаимоисключающих вариантов теории, отсутствие доказательств, неопровержимо свидетельствующих в пользу тихогенеза на всех уровнях эволюции и не могущих быть объясненными никак иначе, противоречия с теорией вероятности, несоответствие с палеонтологической летописью, отсутствие объяснения макроэволюционных изменений и связи структуры генома с эволюцией [Московский, 99], ортогенетические же концепции уделяют слишком мало внимания естественным эволюционным механизмам.
С другой стороны, человек, наделенный даром обобщения и теоретизирования, даже при минимуме информации, а тем более - при достаточно большой фактической подготовке, способен мысленно проследить эволюционные связи между теми или иными группами живых организмов, равно как и создать достаточно убедительную и достоверную теорию по любому другому глобальному вопросу. Примером тому в биологии является, например, предвосхищение клеточной теории умозрительной натурфилософской концепцией Окена или генетической теории наследственности - умозрительной теорией Нэгели об идиоплазме и трофоплазме [Назаров, 91, стр. 60]. Все это вынуждает нас заново пересмотреть глобальные эволюционные представления различных авторов и умозрительно “примирить” друг с другом те из них, которые не находятся в явном противоречии с фактами.
§ 1. 3. ОБЗОР ОСНОВНЫХ ИСТОЧНИКОВ.
Поскольку эволюционная наука является наиболее важной для теоретической биологии, а многие их вопросы до сих пор являются дискуссионными, имеется огромное количество литературы, в той или иной степени затрагивающие вопросы эволюции и систематики, как в целом, так и по отдельным группам. Нет необходимости описывать историю развития эволюционных представлений, так как эта тема знакома практически каждому, поэтому здесь будут отмечены только те литературные источники, которые оказали влияние на формирование наших взглядов по рассматриваемым вопросам.
Прежде всего, к таковым относится монографии Л. Маргелис “Роль симбиоза в эволюции клетки”, В. Жданова “Эволюция вирусов” и В. Назарова “Учение о макроэволюции: на путях к новому синтезу”, статьи И. Мирабдуллаева, опубликованные в журналах “Успехи современной биологии”, “Журнал общей биологии” и “Издание АН СССР, серия биологическая”, статья А. Адрианова и В. Малахова “Сравнительно-морфологический анализ организации головохоботных червей”, опубликованная в “Зоологическом журнале”, а также статья А. Московского “Есть ли научная альтернатива дарвиновской концепции эволюции?” из педагогического вестника “Три ключа”. Из монографии Маргелис и на основе общеизвестных данных была почерпнута идея о симбионтном приобретении эукариотической клеткой отдельных органоидов, что послужило отправной точкой в формировании космическо-теллурической гипотезы происхождения жизни на Земле и симбиогенного возникновении прогенота; в книге Жданова, помимо собственно эволюции вирусов, подробно рассматриваются ранние этапы эволюции жизни; книга Назарова предоставила большой материал и подробный анализ по различным теориям, касающимся вопросов макроэволюции; работы Мирабдуллаева позволили разграничить эукариот на группы по типам митохондриальных крист и другим признакам и составить списки таксонов, входящих в эти группы; статья Адрианова/Малахова позволила предварительно разделить многоклеточных животных на две основные ветви по типу дробления зиготы; в работе Московского мы обнаружили наиболее современный и, по нашему мнению, правдоподобный взгляд на эволюцию жизни как одно из базовых свойств живой материи. Большое значение для нас имели также работы Берга, Беклемишева, Кулаева и некоторых других. Отдельные высказывания и факты, встреченные у некоторых других авторов, также послужили почвой для формирования описанных здесь концепций.
Что касается других источников, то они, в основном, привлекались нами как справочные пособия по различным вопросам, затрагиваемым в ходе работы. Значительное количество информации почерпнуто также из университетских лекций по зоологии беспозвоночных, биологии индивидуального развития, генетике и теории эволюции. Кроме того, при написании этой работы использовалась подборка данных, собранная нами в 1995-96 годах при работе над докладом по теме “Филогенетическое положение мохообразных и родственных им групп в контексте происхождения и эволюции высших растений”, представленным на научно-практической конференции студентов, аспирантов и преподавателей КГУ, а также материалы, собранные для написания контрольной работы по теории эволюции (тема - “Антропогенез”).
1. Современная биология переживает кризис, вызванный, прежде всего, резкой специализацией и математизацией на фоне роста количества фактического материала и нетерпимости между приверженцами различных концепций.
2. Многие новые биологические дисциплины в данный момент не имеют реального значения без выхода на практическое применение или без увязки с эволюционной теоретикой.
3. Современная систематика, претендуя на звание филогенетической, до сих пор сохраняет черты экологической, морфологической и организационно-уровневой систематики.
4. Неустановленность систематических критериев ведет к путанице в систематике разных групп живых организмов.
5. Многие вопросы филогении являются “белыми пятнами” в эволюционной науке.
6. Традиционные методы систематики либо не способны более внести нечто принципиально новое в эволюционную систематику, либо их результаты носят мозаичный характер, в связи с чем назрела необходимость в увязке имеющихся данных в единую картину логическим путем.
7. Тихогенетические (дарвинистские) представления об эволюции доказали свою несостоятельность, тогда как ортогенетические получают все новые подтверждения.
8. Следует заново пересмотреть глобальные исследования крупных эволюционных теоретиков ХХ века и, наложив на них современный фактический материал, обработать их в целях создания новейшей эволюционной теории.
2. 1. МАТЕРИАЛЫ.Поскольку данная работа носит теоретический характер, то можно сказать, что материалом исследования явились литературные источники по исследуемым вопросам. Сбор материалов велся в течение всего времени обучения в Калининградском государственном университете, то есть в период с 1995 по 2000 год, причем часть работы, касающаяся происхождения и эволюции высших растений, докладывалась на Научно-практической конференции студентов, аспирантов и преподавателей КГУ (секция биологическая, подсекция ботаническая) в 1996 году, где заслужила высокую оценку организаторов.
Что касается метода исследования, то он, будучи, фактически, самым древним возможным, не утратил своей актуальности, так как может быть применен к любым областям знаний.
Как и любой другой, умозрительный метод имеет свои плюсы и минусы. К плюсам относится его универсальная применимость к общетеоретическим вопросам (то есть, одним и тем же способом можно действовать при построении теорий в физике, астрономии, истории, теологии, биологии и т. д.), отсутствие абсолютной необходимости конкретных узкоспециальных знаний и навыков (хотя и они, разумеется, могут оказаться полезными), практическая простота (это не значит, однако, что умозрительный метод прост и общедоступен: просто, если человек владеет им, ему не нужны ни оборудование, ни строго заданные условия проведения исследований); наконец, основным плюсом метода является то, что он позволяет сводить имеющиеся факты в единую картину, что недоступно ни одному другому методу. С другой стороны, метод этот бесполезен в практических отраслях науки, не предназначен для прямой, механической передачи другим людям (если результаты умозрительного метода легко довести до окружающих, то для освоения самого метода нужны определенные врожденные или приобретенные еще в раннем возрасте способности), и его результаты могут не согласовываться с какими-либо фактами, неизвестными исследователю или не принятые им в расчет; однако метод обладает особой пластичностью, и при выявлении новых фактов его результаты могут легко изменяться этим или другим исследователем, владеющим методом, без существенных изменений в самой сути теории.
Основной задачей метода является анализ и синтез наибольшего числа фактов и концепций (если последние, разумеется, не находятся в непреодолимом противоречии друг с другом), а не подтверждение или опровержение той или иной конкретной теории. Значение умозрительного (гипотетико-дедуктивного и индуктивного) метода подчеркивается и в работе Назарова [Назаров, 91, стр. 24]. Так, очевидно, что такие важные естественнонаучные методы, как наблюдение и эксперимент, слабо применимы к надвидовой эволюции (макроэволюции в традиционном понимании). Исходя из этого единственный верный путь для всестороннего изучения макроэволюции - реконструкция ее механизма и построение соответствующей теории на основе сопоставления и объединения данных, относящихся к разным уровням биологической организации. При этом важнейшим критерием достоверности умозрительной теории является “требование взаимной логической и фактической непротиворечивости данных” [Назаров, 91, стр. 47-48]. Именно таким положением мы руководствовались, описывая механизмы и результаты эволюции на надвидовом уровне.
.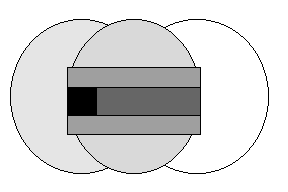
Рис. 3. Положение умозрительного метода эволюционной теории в системе наук.
Умозрительный метод унаследован из гуманитарного крыла фундаментальных наук, так же как статистические - из точного, а, например, наблюдение и эксперимент являются автохтонно естественнонаучными методами. Поскольку биология в частности и естественные науки в целом стоят на пересечении наук точных и гуманитарных (см. рис. 3), в них следует считать допустимыми и равнозначными методы разных направлений, а так как эволюционная теория является гуманитарным, историко-философским направлением биологии, то приоритет в ней, по-видимому, следует отдавать гуманитарным методам, так же, как, например, в биофизике или физиологии - математическим, а в этологии и зоопсихологии - естественным
.Следует отметить также, что этот метод оперирует скорее не с реально произошедшими событиями, а с их вероятностью; однако под вероятностью в данном случае подразумевается не математическая величина, а логическое понятие, определяющее, может ли теоретически произойти то или иное событие. Не исключено, что со временем понятие математической вероятности также будет приложено к данному методу, но в этой работе вероятность будет определяться чисто логическим, умозрительным путем.
1. Главные преимущества умозрительного метода - его универсальная применимость к общетеоретическим вопросам, отсутствие абсолютной необходимости конкретных узкоспециальных знаний и навыков, практическая простота и возможность с его помощью сводить имеющиеся факты в единую картину, что недоступно ни одному другому методу.
2. Главные недостатки метода в том, что он бесполезен в практических отраслях науки, не предназначен для прямой механической передачи другим людям и
его результаты могут не согласовываться с какими-либо фактами, неизвестными исследователю или не принятыми им в расчет.3. Несмотря на ряд недостатков, умозрительный метод не утратил актуальности и до сих пор является перспективным для решения многих эволюционных вопросов.
4. Основной задачей метода является анализ и синтез наибольшего числа фактов и концепций, а не подтверждение или опровержение той или иной конкретной теории.
5. Умозрительный метод зачастую является единственным возможным при рассмотрении макроэволюционных вопросов, так как макроэволюционные изменения в большинстве случаев недоступны для наблюдения и эксперимента, а данный метод оперирует не с реально произошедшими, а с вероятными событиями.
6. Важнейший критерий достоверности результатов в умозрительном методе - взаимная логическая и фактическая непротиворечивость данных.
7. Реальный результат в современной эволюционной науке может дать только тесное взаимодействие между методами точных, естественных и гуманитарных наук.
Глава 3. РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ.
§ 3. 1. ПРИНЦИПЫ УМОЗРИТЕЛЬНОГО МЕТОДА.
Принципами умозрительного метода в эволюционной систематике и теории эволюции (ПУМ) названы здесь некоторые достаточно разрозненные и пока что слабо систематизированные тезисы, касающиеся закономерностей эволюционного процесса, биологической систематизации и действия самого метода, которые следует учитывать при умозрительном анализе теории эволюции и эволюционной систематики для построения наиболее четкой и логически выверенной эволюционной картины. Для начала необходимо отметить некоторые из них (как сформулированные, так и не сформулированные ранее).
ПУМ можно разделить на три основные группы: принципы действия метода (ПДМ), принципы определения (ПО, подразделяющиеся на принципы возможности, возникновения и направленности) и принципы систематизации (ПС) (см. табл. 1) . Следует отметить, что, во-первых, такое разграничение описанных ПУМ достаточно условно, так как один и тот же принцип может иметь разные аспекты и применяться к разным ситуациям, а, во-вторых, что перечень принципов, вероятно, можно расширить. Не исключено также, что в дальнейшем, при более подробной разработке общей теории эволюции, некоторые из перечисленных здесь принципов будут увязаны с положениями о механизмах и движущих силах эволюции и войдут в эволюционную теорию и систематику, а также в общебиологическую методологию.
3. 1. 2. Принципы действия метода.
К принципам действия метода относятся принципы мозаичности, презумпции позитивизма, исключений и лабильности.
Принцип мозаичности. Любая полнота фактического материала недостаточна для полного прослеживания эволюционных связей. Исходя из этого принципа, вне зависимости от имеющихся данных (прежде всего палеонтологических, так как многие эволюционные преобразования могли не оставить никакого следа в палеонтологической летописи, и биохимических, так как с точки зрения теории вероятности выявить все биохимические особенности всех ныне живущих и тем более вымерших видов невозможно) для эволюциониста-систематика необходимо умение создавать умозрительные построения.
Таблица 1. Принципы Умозрительного Метода.
|
Принципы Умозрительного Метода |
Принципы действия метода |
|
Принцип мозаичности |
|
|
|
|
Принцип презумпции позитивизма |
|
|
|
|
Принцип исключений |
|
|
|
|
Принцип лабильности |
|
|
Принципы определения |
Принципы возможности |
Принцип вероятности |
|
|
|
|
Принцип комбинирования |
|
|
|
Принципы возникновения |
Принцип парафилетичности |
|
|
|
|
Принцип вариабельности |
|
|
|
|
Принцип тупиковости специализации |
|
|
|
|
Принцип филогенетического ускорения |
|
|
|
Принципы направления |
Принцип экономичности развития |
|
|
|
|
Принцип изъятия лишнего |
|
|
|
|
Принцип экологического давления |
|
|
|
|
Принцип эволюционных тенденций |
|
|
|
|
Принцип холизма |
|
|
|
|
Принцип направленности |
|
|
|
|
Принцип допустимости |
|
|
|
|
Принцип сопряженности |
|
|
Принципы систематизации |
|
Принцип детерминирования |
|
|
|
|
Принцип кладирования |
|
|
|
|
Принцип унификации |
|
|
|
|
Принцип повышения статуса |
Принцип презумпции позитивизма
таков: если какое-либо утверждение не опровергнуто и не доказано, то его следует считать возможным. Исходя из этого принципа, все механизмы, которые когда-либо предполагались в качестве движущих сил эволюции и не были достоверно опровергнуты, можно рассматривать как реально имевшие или имеющие место в эволюционном процессе.Принцип исключений можно афористически сформулировать следующим образом: если закон подтверждается исключениями, то он плохо сформулирован. Другими словами, всякое имеющее место исключение должно восприниматься не как подтверждение правила, а именно как исключение, и появление каждого нового исключения из старого правила, каким бы незначительным оно ни казалось, требует изменения правила.
Наконец, важен в умозрительных построениях принцип лабильности, согласно которому всякие новые данные могут быть уложены в прежнее построение умозрительным же способом.
3. 1. 3. Принципы определения.
Принципы определения подразделяются на принципы возможности, принципы возникновения и принципы направления в зависимости от того, какую часть эволюционного процесса они затрагивают.
К принципам возможности относятся те ПУМ, которые рассматривают вероятности появления тех или иных признаков у разных групп живых организмов. Нами сформулированы два принципа этой группы: принцип вероятности и принцип комбинирования.
Согласно принципу вероятности, для установления родственных связей между видами и группами, обладающими сходными структурными, экологическими, физиологическими и другими признаками, следует выяснить, насколько вероятно их моно- или полифилетическое возникновение. При этом очевидно, что чем сложнее признак, чем из большего числа составляющих он состоит (или же чем больше сходных признаков у данных видов), тем больше вероятность, что группа, им обладающая, является монофилетической (в широком смысле слова, включающем также понятие “парафилетический”); обратная формулировка при этом не верна: простой признак может возникнуть как поли-, так и монофилетически. Так, высока вероятность того, что жгутик эукариот, обладающий сложной упорядоченной структурой 9+Х (где Х может быть равно 0, 1 или 2), возник монофилетически (в данном случае не рассматривается возможность его полифилетичного приобретения путем симбиоза согласно гипотезе Линн Маргелис: возникла эта структура, в любом случае, скорее всего, монофилетично, пусть и у гипотетических прокариотических симбионтов [Маргелис, 83]), тогда как простой флагеллиновый жгутик прокариот мог возникнуть как один, так и несколько раз независимо.
Принцип комбинирования гласит, что если какой-то признак мог возникнуть в одной группе, то существует ненулевая вероятность его возникновения в другой, причем, с одной стороны (исходя из принципа вероятности), чем сложнее признак, тем эта вероятность меньше, а с другой (исходя из принципа эволюционных тенденций) – чем группы эволюционно ближе друг к другу, тем она больше. Из этого ПУМ также следует, что возможно существование “химерной” группы, сочетающей признаки других групп, но вероятность существования такой группы уменьшается при увеличении числа и сложности этих признаков, а также при удаленности друг от друга и от гипотетической “химеры” реальных носителей этих признаков. Принцип этот следует учитывать при умозрительных эволюционных построениях.
Принципами возникновения называются ПУМ, оперирующие с базовыми группами, в которых происходит возникновение и накопление признаков. К ним относятся принципы парафилетичности, вариабельности, тупиковости специализации и филогенетического ускорения.
Согласно принципу парафилетичности, монофилетичность примитивного признака не свидетельствует о монофилетичности прогрессивного признака того же эволюционного ряда. Чем древнее признак, тем больше вероятность его монофилетичности: группа, обладающая примитивным признаком, скорее всего, является базовой, парафилетической, а от нее моно- или полифилетично (причем под монофилией может подразумеваться не только происхождение от единственной особи или даже популяции, но и от вида в целом, при использовании идентичных путей видообразования в разных популяциях) отделяются группы с одинаковыми или различными прогрессивными признаками. Примером здесь может являться конечность млекопитающих: млекопитающие с пятипалой стопоходящей конечностью составляют парафилетический пласт, от которого одним или разными путями отпочковываются ветви, ведущие к копытному пальцехождению через некопытное пальцехождение (к конечности, подобной лошадиной, через конечность, подобную собачьей; причем, вероятнее всего, такая эволюция полифилетично привела к возникновению разных групп копытных), а также к многочисленным другим модификациям лапы [Наумов, Карташев,79, стр. 207-208].
Принцип вариабельности заключается в том, что в древних группах признаки могут быть не устоявшимися, то есть даже близкородственные виды могут иметь существенные различия, на более высоких этажах эволюции считающиеся систематически значимыми. Так, у кишечнополостных обнаружены все основные типы дробления: от примитивного хаотического до примитивных форм спирального и радиального [Догель, 80; Захваткин, 49], - тогда как у более прогрессивных групп характер дробления лежит в основе разделения на две большие ветви - Spiralia и Radialia [Адрианов, Малахов, 85]. Это говорит о том, что в примитивных группах может формироваться определенный фонд для дальнейших преобразований, из которого берут начало прогрессивные ветви.
С принципом вариабельности связан также принцип тупиковости специализации. Согласно этому принципу, чем более специализирован организм, тем меньше вероятность его эволюционирования (кроме микроэволюции, не зависящей от степени специализации [Грант, 80, 91]). Действительно, вид, специализирующийся, допустим, на строго определенной пище (стенотроф), гибнет при ее отсутствии, а не перестраивается на другую пищу; крыло летучей мыши, копыто оленя или ласт моржа также не может эволюционировать ни во что другое [Жизнь - как она возникла? 85]. Этот ПУМ является одним из наиболее важных для осознания изложенных ниже представлений о механизмах эволюционного процесса, так как, с традиционной точки зрения, полностью противоречит самому факту эволюции, поскольку большинство современных видов, если не все, в чем-либо специализированы. Однако это отнюдь не свидетельствует о том, что эволюция в данный момент прекратилась или не шла вовсе: просто подходить к ней нужно с совершенно других позиций. Для разъяснения этого принципа следует отметить также, например, тот факт, что приматы как отряд с биологической точки зрения довольно примитивны, что и дало им возможность для эволюционного скачка в плане развития ЦНС. Более же подробно вопросы, связанные с этим ПУМ, будут освещены во второй части работы.
Принцип филогенетического ускорения общеизвестен. Он сформулирован Бергом и заключается в том, что “индивидуальное развитие может не только повторять филогению, но и предварять ее” [Берг, 77, стр. 88]
Принципы направления (принципы экономичности развития, изъятия лишнего, экологического давления, эволюционных тенденций, холизма, допустимости, направленности и сопряженности) связаны с механизмами, направляющими эволюцию.
Согласно принципу экономичности развития, эволюция идет преимущественно по наименее энергоемкому пути. Из этого принципа вытекает и положение о необратимости эволюции [Кейлоу, 86] (не применимый, однако, к процессам микроэволюции): абсолютно неэкономичен (и потому нереален) выход на определенную эволюционную ступень с последующим возвратом на прежний уровень (упрощение организмов под воздействием прикрепленного или паразитического образа жизни, возврат в водную среду или, например, вторичный переход обезьян-панид к четвероногому способу передвижения от распрямленных двуногих предков - не эволюционный возврат, а крайние формы прогрессивной эволюции под воздействием специфических условий среды; иными словами, катагенез есть видоизмененная форма арогенеза). Так, не логично предполагать, что выделяющие мочевину млекопитающие происходят от выделяющих мочевую кислоту рептилий, происходящих, в свою очередь, от выделяющих мочевину амфибий [Наумов, Карташев, 79, т. 2, стр. 46-47]: гораздо логичнее выглядят предположения о независимом происхождении рептилий и млекопитающих от разных групп амфибий либо даже о происхождении амфибий, рептилий и млекопитающих (или, что вероятнее, различных групп “амфибоидов”, а также анапсидных, диапсидных и синапсидных позвоночных) от разных групп рипидистиевых кистеперов. Примечательно, что к аналогичному выводу приводит нас и сравнение суставов конечностей амниот, где одну из групп образуют современные амфибии и млекопитающие, а другую - рептилии и птицы [Наумов, Карташев, 79, т. 2, стр. 207].
Близок к принципу экономичности принцип изъятия лишнего, согласно которому вызывающие энергетическую неэкономичность органы, процессы или стадии развития утрачиваются или упрощаются до минимально необходимых. Таким образом, виды, обладающие “балластными” признаками, от которых теоретически можно избавиться без вреда для организма, скорее всего, являются более примитивными, чем те, у которых ничего лишнего нет. На основе этого принципа можно, например, выстроить эволюционный ряд изменения процесса гаструляции: мультиполярная иммиграция - униполярная иммиграция - плотное врастание - инвагинация, - так как движение клеточных пластов более совершенно и экономично, нежели несогласованное движение клеток [Беклемишев, 79, стр. 38; Богомолов, 68; Захваткин, 49; Зусман, 73; Иванов, 68].
Принцип экологического давления в какой-то степени противоположен принципу изъятия лишнего и заключается в том, что наиболее долго в неизменном виде сохраняется тот признак, который менее всего испытывает давление окружающей среды. Согласно этому принципу, наиболее долго сохраняются именно признаки, не играющие существенной адаптивной роли, однако признаки эти обычно мало обращают на себя внимание. Хорошим примером этого может служить удивительное сходство жилкования крыльев верблюдок и златоглазок, иногда разносимых по разным отрядам. Поскольку характер жилкования, а тем более, небольшие его детали, если и влияет на аэродинамику крыла, то не особо значительно (ведь существует огромное количество типов жилкования даже у видов, летающих сходным образом), а также исходя из принципа вероятности, можно сделать вывод, что параллельное развитие практически идентичного жилкования не только в разных отрядах, но и в разных семействах, а то и родах, невозможно, и, следовательно, что верблюдки и златоглазки являются ближайшими родственниками. Три описанных выше принципа можно сформулировать в виде одного афоризма: “Лень - двигатель эволюции”.
Интересен также принцип эволюционных тенденций, согласно которому моно- либо полифилетически может возникать не только какой-либо конкретный признак, но и тенденция к его возникновению (эволюционную тенденцию можно, таким образом, рассматривать как самостоятельный биологический признак), причем, чем ближе филогенетически находятся таксоны, тем больше сходство в их эволюционных тенденциях. Механизмы такой “эволюции тенденций” остаются неясными (возможно, они кроются в “спящих генах”, может быть, дело в закономерностях между планом строения и дальнейшей эволюцией [Северцев, 90], а может быть, тут не обошлось без некой “Высшей Силы”), но примеры этого многочисленны, на что обратил внимание еще Л. Берг в теории номогенеза [Грант, 80]. Так, эволюция Lamellicristata (организмов с пластинчатыми кристами митохондрий) наблюдаются тенденции к многоклеточности (животные, растения, грибы, багрянки; за пределами Lamellicristata - только бурые водоросли из Tubulicristata), к выходу на сушу (животные - неоднократно, растения, грибы; за пределами группы - только различные слизевики, предпочитающие, однако, влажные места) [Мирабдуллаев], хотя сами эти признаки (многоклеточность и сухопутность), несомненно, полифилетичны. Подобным образом архозавры имели тенденцию к двуногости (даже крокодилы и гигантские четвероногие динозавры, по некоторым данным, являются четвероногими вторично) и к полету (птерозавры, птицы) [Наумов, Карташев,79, т. 2, стр. 13-14].
Принцип холизма заключается в том, что целое есть не сумма составляющих его частей, а другая форма организации материи. Иными словами, элементарные частицы, атомы, молекулы, клетки и живые организмы есть не разные уровни организации, а всего лишь количественно различные, но равные качественно формы (холоны). Исходя из этого принципа, можно сделать вывод, что эволюция неорганического и органического мира основывается на одних и тех же законах, модифицированных для данных объектов [Московский, 99].
Принцип направленности заключается в том, что система живых организмов существует объективно, а эволюция только заполняет имеющиеся в ней вакансии [Московский, 99].
Из принципа направленности вытекает и принцип допустимости, который говорит о том, что эволюционные изменения происходят в некоем допустимом вероятностном пространстве.
К принципам направления относится также принцип сопряженности, согласно которому эволюционные преимущества имеют не те организмы, которые побеждают в борьбе за существование, а те, которые наиболее способствуют эволюции Вселенной в целом. Сопряженность эволюции связана с предопределенностью и объективностью системы живых организмов [Вернадский, 60; Молчан, 98].
3. 1. 4. Принципы систематизации.
В отличие от ПО, описывающих действие самого эволюционного процесса, и ПДМ, пригодных для применения метода к любому материалу (а не только биологическому), ПС служат для составления удобной и филогенетически верной системы. К принципам систематизации здесь отнесены принципы детерминирования, кладирования, унификации и повышения статуса.
Принцип детерминирования заключается в том, что чем раньше в филогенезе или, соответственно, в онтогенезе закладывается тот или иной признак, тем большее систематическое значение он имеет. Исходя из этого принципа, наиболее важно в классификации животных строение сперматозоидов и яйцеклеток, далее идет характер дробления, далее - тип гаструляции, способ закладки целомов и т.д. [Адрианов, Малахов, 85; Беклемишев, 79; Захваткин, 49; Иванов, 68; Малахов, Реунов].
С точки зрения принципа кладирования, новая систематическая группа возникает не с достижением предковой группой определенного уровня развития, свойственного этой новой группе, а с появлением предпосылок к возникновению характерной для данной группы организации [Павлинов, 90]. Пользуясь этим принципом, можно избежать типичной для современной систематики проблемы, когда, с одной стороны, трудно разделить ископаемые формы между предковой и потомковой группой, так как одни возможные предки близки к потомкам по одному признаку, а другие - по другому, а с другой - избежать полифилетичности и, в какой-то мере, парафилетичности таксона. Характерный пример: при выделении группы Mammalia сомнительно систематическое положение некоторых тероморф, тогда как при выделении монофилетичной группы Sinapsida эта проблема отпадает.
Принцип унификации вытекает из самой идеи единства живой природы и предполагает, что к любым систематическим группам применимы одни и те же критерии классифицирования, хотя не для всех групп эти критерии найдены. Из этого принципа, в частности, следует необходимость согласования зоологический и ботанической классификации и таксономии (или, если быть точнее, необходимость создания единой биологической систематики и таксономии), а также потребность в оценке значимости признаков (что можно сделать, основываясь, например, на других ПС, а также с учетом механизмов эволюционных преобразований).
Принцип унификации следует согласовывать с принципом повышения статуса, гласящим, что с повышением уровня организации повышается общая значимость систематических признаков. Так, в основе деления на царства наиболее примитивной (и, возможно, наиболее древней) группы живых организмов - вирусов - лежит такой глобальный признак, как характер наследственного материала и стратегия его реализации [Жданов,66, стр. 49], тогда как у клеточных форм жизни этот признак един для всех (наследственность обусловлена двойной спиралью хромосомной или хромонемной ДНК, причем информация реализуется с помощью т- и иРНК), а деление на царства происходит на другом основании; выделение типов внутри прокариотических царств основывается на особенностях клеточного метаболизма, тогда как, например, внутри царств Zoobionta и Phytobionta метаболизм более-менее однообразен.
Приведенные выше ПУМ (хотя перечень их, по-видимому, не исчерпывающ) являются не только основными принципами умозрительного подхода к теории эволюции, но и (особенно ПО) важными эволюционными принципами. Основываясь на ПУМ и имеющихся у нас в распоряжении фактах, можно выделить основные эволюционные механизмы и выстроить возможную картину эволюции в живой природе. При этом следует учитывать соответствие различных гипотез как можно большему числу описанных ПУМ.
1. Для описания различных сторон эволюционного процесса, эволюционной систематики и умозрительного метода можно выделить некоторые принципы умозрительного метода (ПУМ), которые следует учитывать при умозрительном анализе теории эволюции и эволюционной систематики для построения наиболее четкой и логически выверенной эволюционной картины.
2. ПУМ можно разделить на принципы действия метода (ПДМ), принципы определения (ПО, подразделяющиеся на принципы возможности, возникновения и направленности) и принципы систематизации (ПС): к ПДМ относятся принципы мозаичности, презумпции позитивизма, исключений и лабильности, к принципам возможности - принцип вероятности и принцип комбинирования, к принципам возникновения - принципы парафилетичности, вариабельности, тупиковости специализации и филогенетического ускорения, к принципам направления - принципы экономичности развития, изъятия лишнего, экологического давления, эволюционных тенденций, холизма, допустимости, направленности и сопряженности, к ПС - принципы детерминирования, кладирования, унификации и повышения статуса.
3. ПУМ являются не только основными принципами умозрительного подхода к теории эволюции, но и (особенно ПО) важными эволюционными принципами.
4. Любая полнота фактического материала недостаточна для полного прослеживания эволюционных связей.
5. Если какое-либо утверждение не опровергнуто и не доказано, то его следует считать возможным.
6. Если закон подтверждается исключениями, то он плохо сформулирован; иначе - всякое имеющее место исключение должно восприниматься не как подтверждение правила, а именно как исключение, и появление каждого нового исключения из старого правила, как бы незначительно оно ни казалось, требует изменения правила.
7. Всякие новые данные могут быть уложены в прежнее построение умозрительным же способом.
8. Для установления родственных связей между видами и группами, обладающими сходными признаками, следует выяснить, насколько вероятно их моно- или полифилетическое возникновение; чем сложнее признак, чем из большего числа составляющих он состоит (или же чем больше сходных признаков у данных видов), тем больше вероятность, что группа, им обладающая, является монофилетической; простой признак может возникнуть как поли-, так и монофилетически.
9. Если какой-то признак мог возникнуть в одной группе, то существует ненулевая вероятность его возникновения в другой; чем сложнее признак, тем эта вероятность меньше; чем группы эволюционно ближе друг к другу, тем она больше.
10. Монофилетичность (в том числе парафилетичность) примитивного признака не свидетельствует о монофилетичности прогрессивного признака того же эволюционного ряда.
11. В древних группах признаки могут быть не устоявшимися.
12. Чем более специализирован организм, тем меньше вероятность его эволюционирования.
13. Индивидуальное развитие может не только повторять филогению, но и предварять ее.
14. Эволюция идет преимущественно по наименее энергоемкому пути.
15. Катагенез есть видоизмененная форма арогенеза.
16. Вызывающие энергетическую неэкономичность органы, процессы или стадии развития утрачиваются или упрощаются до минимально необходимых.
17. Наиболее долго в неизменном виде сохраняется тот признак, который менее всего испытывает давление окружающей среды.
18. Моно- либо полифилетически может возникать не только какой-либо конкретный признак, но и тенденция к его возникновению.
19. Целое есть не сумма составляющих его частей, а другая форма организации материи.
20. Система живых организмов существует объективно, а эволюция только заполняет имеющиеся в ней вакансии.
21. Эволюционные изменения происходят в некоем допустимом вероятностном пространстве.
22. Эволюционные преимущества имеют не те организмы, которые побеждают в борьбе за существование, а те, которые наиболее способствуют эволюции Вселенной в целом.
23. Чем раньше в филогенезе или, соответственно, в онтогенезе закладывается тот или иной признак, тем большее систематическое значение он имеет.
24. Новая систематическая группа возникает не с достижением предковой группой определенного уровня развития, свойственного этой новой группе, а с появлением предпосылок к возникновению характерной для данной группы организации.
25. К любым систематическим группам применимы одни и те же критерии классифицирования, хотя не для всех групп эти критерии найдены.
26. С повышением уровня организации повышается общая значимость систематических признаков.
27. Основываясь на ПУМ и имеющихся у нас в распоряжении фактах, можно выделить основные эволюционные механизмы и выстроить возможную картину эволюции в живой природе.
§ 3. 2. УМОЗРИТЕЛЬНЫЙ АНАЛИЗ ТЕОРИИ ЭВОЛЮЦИИ:
УРОВНИ И МЕХАНИЗМЫ ЭВОЛЮЦИИ.
В настоящее время принято разделять эволюционный процесс на два уровня - микро- и макроэволюцию; при этом микроэволюцией обычно называется эволюция на уровне популяций, ведущая к видообразованию, а макроэволюцией - эволюция на надвидовом уровне [Грант, 80, 91; Назаров, 91; Черникова, 87]. Однако это деление в значительной степени условно, и, прежде всего в связи с тем, что до сих пор не существует точного и всеобъемлющего определения вида (все существующие определения имеют ряд ограничений: например, для видов без полового размножения, видов-двойников, видов с кольцевыми ареалами и т.п. [Грант, 80]; попытка универсального определения видов по установлению мономорфных видоспецифичных участков генома [Назаров, 91, стр. 245] пока что не получила практического распространения), а тем более - крупных таксонов. Поэтому, вероятно, следует разграничить уровни эволюции, основываясь на других принципах, нежели на заведомо условных таксономических, например, на эволюционных механизмах, характерных для того или иного уровня. В пользу такого предположения говорит и гипотеза Мейена о предопределенности эволюции и независимости систематики от количества исследованных групп [Московский, 99].
Имеется также и другая концепция подразделения эволюционного процесса: по уровням биологической организации, затрагиваемым данными уровнями преобразования живой материи. Основываясь на таком принципе, историческое развитие жизни подразделяется на онтогенез (изменения на организменном уровне), филогенез, включающий микроэволюцию (изменения на популяционно-видовом уровне) и макроэволюцию (изменения на уровне видов и выше, с момента возникновения генетической изоляции), и сингенез (изменения на биосферно-биогеоценотическом уровне) [Назаров, 91, стр. 11-12]. Процессы масштабом ниже онтогенетического попадают в компетенцию физиологии, биохимии, генетики и т. п., а выше сингенетического - в компетенцию астрономических наук (см. табл. 3). Общая же теория эволюции, создание которой планируется в результате синтеза современных эволюционных концепций, должна затрагивать и онто-, и фило-, и сингенез, и даже пограничные области соседствующих с ними в этой системе дисциплин, касающихся, в основном, циклических процессов развития.
Таблица 3. Сфера компетенции общей теории эволюции.
|
Этап эволюционирования |
Сфера компетенции |
Науки |
|
космогенез |
космические процессы |
астрономия |
|
сингенез |
историческое развитие сообществ и биосферы |
эволюционная экология |
|
филогенез |
историческое развитие живой природы |
теория эволюции |
|
онтогенез |
индивидуальное развитие организмов |
эмбриология, биология постнатального онтогенеза |
|
|
жизнедеятельность организмов |
физиология, генетика, биохимия и т. п. |
По мотивам, которые будут понятны из дальнейшего изложения, филогенез разделен нами на три уровня: микро-, мезо- и макроэволюция (или, во избежание омонимии, мегаэволюция), причем их границы несколько отличаются от традиционно принятых (см. рис. 4) и не совпадают с границами тех или иных традиционно выделяемых систематических групп: микроэволюция, имеющая в своей основе "дарвиновские", тихогенетические механизмы эволюции [Дарвин, 52], затрагивает внутривидовые таксоны, виды, а в крайнем случае - роды; мезоэволюция, основой которой являются макромутации (прежде всего, связанные с изменением числа хромосом - как кратного, так и некратного), мутации регуляторных генов, приводящие к гетеротопиям и гетерохрониям типа акцелерации, гиперморфоза, прогенеза и неотении [Назаров, 91, 205] и горизонтальный перенос подвижных элементов ДНК [Тихоненко, 80; Гвоздев, 98; Глазер, 98; Назаров, 91, стр. 41; Хесин, 84], влияют на образование новых родов, семейств, отрядов и групп более высокого порядка (возможно, иногда - до типа включительно); наконец, мегаэволюция, механизмы которой связаны с изменением энергидности и степени самостоятельности субъектов биологических конгломератов, ведет к образованию типов,
царств и более крупных групп и основывается на таких явлениях, как колониализация, межвидовая гибридизация, симбиогенез, автономизация клеточных элементов и т. п. (см. табл. 3). Элементарным процессом микроэволюции являются мутации и рекомбинации, мезоэволюции - генетическая изоляция (как первичная, так и вторичная), а мегаэволюции - мегаэволюционная детонация, явление, которое будет описано в дальнейшем. Микроэволюция связана с изменением генетической структуры, мезоэволюция - с преобразованием общей схемы организации [Назаров, 91, стр. 14], а мегаэволюция - с изменением энергидности. Помимо этих трех уровней филогенеза, будут рассматриваться также некоторые вопросы, касающиеся сингенеза в планетарном и космическом масштабе.Таблица 3. Уровни эволюции.
|
Уровни |
микроэволюция |
мезоэволюция |
мегаэволюция |
|
Таксоны |
популяции, сорта, породы, виды, роды |
роды, семейства, отряды, классы, типы |
типы, царства и выше |
|
Изменения |
медленные, постепенные, случайные |
мгновенные, случайные |
изначально запрограммированные, мгновенные, но с постепенным повышением уровня облигатности |
|
Механизмы |
микромутации структурных генов поток генов дрейф генов борьба за существование естественный отбор |
макромутации (в том числе полиплоидия и анеуплоидия) мутации регуляторных генов, ведущие к гетерохрониям и гетеротопиям горизонтальный перенос генов |
колониализация симбиогенез межвидовая гибридизация автономизация клеточных структур |
|
Направление |
ненаправленный процесс |
аллогенез |
арогенез, катагенез |
|
Результаты |
разнообразие родственных форм |
генетическая изоляция |
новые формы и уровни организации жизни |
|
Ограничения |
допустимые пределы вариаций вида |
летальность большинства макромутаций? |
биологическая целесообразность |
Как можно заметить, "мишени" различных уровней эволюции в какой-то степени пересекаются друг с другом. Это связано, однако, не с тем, что на одном и том же уровне действуют разные механизмы, а со значительной условностью современной систематики, в которой аналогичным образом возникшие группы могут иметь разный таксономический статус. Что интересно, эти три уровня эволюции, по-видимому, не пересекаются: вопреки традиционному мнению, мезоэволюция и мегаэволюция отнюдь не складываются из множества микроэволюционных изменений, а протекают совершенно независимо [Четвериков, 26; Назаров, 91, стр. 30]. Именно такой вывод можно сделать из множества фактов, когда тот или иной орган имеет полезные свойства только во всей своей сложности и, следовательно, не мог возникнуть путем постепенного усовершенствования. Примером тому могут служить, например, крыло птиц или насекомых, глаз позвоночных и многие другие признаки, для объяснения которых просто необходимо найти подходящую естественнонаучную концепцию, дабы не давать лишнего преимущества сторонникам творения [Жизнь - как она возникла? 85, стр. 71-82; Московский, 99].
А В
Рис. 4. Соотношение традиционного (А) и принятого в данной работе (В) представления об уровнях эволюционного процесса.
3. 2. 2. Особенности микроэволюции.
Механизмы, отнесенные в данной работе к механизмам микро- и мезоэволюции, хорошо известны в теории эволюции. Однако если механизмы микроэволюции (микромутации, рекомбинации, борьба за существование, естественный отбор, поток генов и дрейф генов) и по традиционным учениям относились именно к микроэволюции и видообразованию [Грант, 80, 91], то механизмы, отнесенные здесь к мезоэволюции, были разделены между процессом видообразования и макроэволюции.
Поскольку учение о микроэволюции не слишком сильно изменилось со времен Дарвина, а ее механизмы хорошо изучены теоретически и практически (например, путем сравнения родственных видов-эндемиков или видов с кольцевыми ареалами наподобие Larus argentatus-fuscus), здесь микроэволюция рассматриваться не будет (кроме того, целью работы является выявление эволюционных связей в более крупных масштабах, нежели масштабы вида и рода). Здесь же следует только дать определение микроэволюции как процесса постепенного ненаправленного преобразования видов. При этом следует отграничивать микромутации как механизм, формирующий микроэволюционные изменения (новые аллели), и рекомбинации, как механизм, образующий новые их комбинации, от других общеизвестных механизмов, регулирующих лишь соотношение аллелей в популяции. Эти механизмы можно объединить под общим названием движущие силы микроэволюции.
Следует отметить, что, в отличие от эволюции более крупных уровней, микроэволюция является обратимым процессом. Действительно, точковые микромутации, не ведущие к резкому изменению выживаемости и размножаемости, возникают случайно, и так же случайно могут возникнуть мутации, им противоположные (прямые и обратные транслокации и инверсии, дупликации - делеции и т. п.); борьба за существование и естественный отбор, хотя и не случайны по своей природе, ведут только к относительной приспособленности к данным условиям, поэтому при случайном изменении условий сначала в прямом, а затем в обратном направлении, тоже будут возвращать популяцию к исходным формам; поток генов и дрейф генов - случайные явления, к тому же, поток генов ведет к изменению соотношения аллелей в пределах популяции, а не вида. Иногда говорят даже, что микроуровень эволюции вообще является не собственно эволюцией, как мезо- и мегауровень, а адаптивной внутривидовой дифференцировкой [Назаров, 91, стр. 41].
Впрочем, микроэволюция тоже может вести к изоляции вида, однако изначально это будет обычно не генетическая, а какая-либо другая форма изоляции (географическая, экологическая и т. п.). Подобным способом, например, возникли уже упомянутые L. argentatus и L. fuscus [Назаров, 91, стр. 29-30]. Таким образом, полностью отрицать видообразовательное (и макроэволюционное в традиционном понимании) значение микроэволюционных процессов мы, безусловно, не можем. Тем не менее, нам представляется более распространенным сальтационное (мезоэволюционное) видообразование с первичностью генетической изоляции.
3. 2. 3. Понятие и механизмы мезоэволюции.
Мезоэволюцию можно охарактеризовать как эволюционный процесс, ведущий ко внезапной (в течение одного поколения) генетической изоляции без предварительного отбора. Как уже было сказано, основными механизмами мезоэволюции являются макромутации, мутации регуляторных генов и горизонтальный перенос генов.
Одним из наиболее часто встречаемых (в основном, среди растений) способов возникновения систематических групп мезоэволюционного уровня является полиплоидия, ведущая к мгновенному (без предварительного накопления каких-либо “полезных признаков”, что еще раз свидетельствует об отграниченности мезоэволюционного процесса от микроэволюционного) возникновению репродуктивной изоляции и, соответственно, к образованию нового вида [Эволюция генома, 86] (возможно, более точно следует считать это событие возникновением нового рода, так как здесь будет отсутствовать даже возможность появления стерильных гибридов с предковой формой, имеющее место при скрещивании видов одного рода, как в случае с родом Equus). Впрочем, полиплоидизацию можно отнести и к мегаэволюции, которая по определению связана с изменением энергидности.
Помимо полиплоидии, изменение числа хромосом, ведущее к видообразованию, может происходить и другими путями. Так, может произойти удвоение не всего набора хромосом, а только части хромосом (анеуплоидия); кроме того, может иметь место утрата одной или нескольких хромосом, разделение одной хромосомы на несколько или слияние нескольких в одну в результате неправильно протекшего процесса митоза. В большинстве случаев, разумеется, такие изменения в генотипе являются летальными или ведущими к стерильности, однако несомненно, что иногда подобные преобразования могут привести и к возникновению новых видов. Так, по-видимому, возникновение разных видов приматов гоминидно-панидной ветви, имеющих разное число хромосом, но поразительное сходство в синтезируемых белках, связано именно со случайными изменениями числа хромосом [Назаров, 91, стр. 40].
Фактическая гомологичность хромосом между собой и с витоспорами (см. п. 3. 3. 1.) свидетельствует о том, что и поли-, и анеуплоидию, и объединение или разделение хромосом можно отнести и к мегаэволюционным механизмам, тогда как не особенно крупные по масштабам воздействия на генотип мутации регуляторных генов могут привести к явно мезоэволюционным преобразованиям. Наиболее существенно в этом плане явление гетерохронии (в частности, неотения), происходящее в результате мутаций генов, регулирующих сроки полового созревания. Хотя генетическая перестройка при неотении небольшая, она позволяет “сбросить груз специализации” и, таким образом, снимает действие принципа тупиковости специализации, направляя эволюцию в горизонтальном (неотеническом) и вертикальном направлении (см. рис. 5); наиболее ярким примером этого процесса мы можем считать эволюцию животных. Кроме того, неотения приводит к мгновенной репродуктивной изоляции, хотя в некоторых случаях (классический пример - амблистома и аксолотль) неотения может быть факультативной. Гетерохронии типа прогенеза и неотении ведут к педоморфозу, а гетерохронии типа акцелерации и гиперморфоза - к рекапитуляциям [Назаров, 91, стр. 205]
. Гетеротопии, в отличие от гетерохроний, ведут к изменению не времени, а места закладки органов, либо к изменению их пространственной ориентации. Как и гетерохронии, гетеротопические изменения сыграли существенную роль в эволюции высших таксонов животных.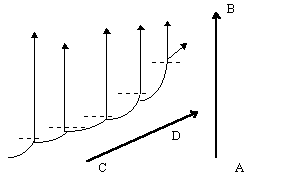
Рис. 5. Соотношение вертикальной и горизонтальной (диагональной) эволюции:
пунктиром отмечено неотеническое “срезание” поздних этапов онто- и филогенеза; треугольные стрелки - прогрессивная эволюция в филумах; вектор AB - вертикальная эволюция; вектор CD - горизонтальная (диагональная) эволюция.
Интересным и своеобразным механизмом мезоэволюции могут являться так называемые
онтомутации - мутации материнской иРНК, контролирующей дробление яйцеклетки и начальные стадии онтогенеза. Поскольку онтомутации возникают у зародыша на начальных стадиях развития, они могут не вести к тяжелым последствиям для организма и одновременно способны в течение одного поколения резко изменить план его строения. Кроме того, гипотеза об онтомутациях снимает вопрос о происхождении нового таксона от единственного “обнадеживающего урода”, так как схожие мутации иРНК, в отличие от мутаций в генотипе, могли бы возникать у значительной части популяции под влиянием одного и того же фактора [Назаров, 91, стр. 151-157].Все макромутационные изменения могут происходить на генах, возникших в результате дупликации, в том числе множественной, что не только снижает вероятность их летальности, но и ведет к общему увеличению содержания ДНК, а также к возникновению принципиально новых генетических локусов, и может способствовать прогрессивной эволюции [Назаров, 91, стр. 236]. Тот же результат дают, помимо дупликации генов, и другие способы увеличения количества генетического материала, такие, как уже упомянутые поли- и анеуплоидия.
В качестве механизма мезоэволюции можно указать также явление горизонтального переноса подвижных участков ДНК с помощью вирусов (вирусная трансдукция) и другими способами (с помощью эпигенов, транспозонов и мобильных генетических элементов [Назаров, 91, стр. 41; Хесин, 84]). Его особенностью является обмен генетической информацией между любыми видами живых организмов без каких-либо ограничений степенью родства [Тихоненко, 80; Гвоздев, 98; Глазер, 98; Жданов, 66, стр. 42].
Важным для мезоэволюции явлением (не механизмом мезоэволюции, а именно явлением, с ней связанным) можно назвать преадаптации, которые могут возникать либо как второстепенные признаки, сопутствующие полезным преобразованиям, либо в результате мутаций, не ведущих к изменению выживаемости в момент их возникновения, но приводящих к улучшению приспособленности в дальнейшем, либо в результате высвобождения органа от ранее выполняемой функции [Назаров, 91, стр. 74]. Примеры таких преадаптаций можно найти опять-таки среди приматов. Так, в результате обеднения ландшафтов перейдя от брахиации к наземному прямохождению, предки человека сохранили вертикальное положение тела, что позволило им легче приспособиться к условиям открытого пространства. Хватательная конечность, сформировавшаяся у приматов для облегчения передвижения по деревьям, при переходе к наземному образу жизни освободилась для выполнения других нагрузок. Таким образом, преадаптациями можно назвать всякие признаки, не возникающие в результате изменения внешних условий, а помогающие к ним приспособиться, приобретая новые функции [Георгиевский, 74], что опять-таки подчеркивает, что микроэволюционные законы мало уместны для описания мезоэволюции.
3. 2. 4. Понятие и механизмы мегаэволюции.
Термин “мегаэволюция” был предложен Дж. Симпсоном для обозначения наиболее крупномасштабных преобразований, связанных с возникновением высших таксонов, начиная с семейства или отряда, но позднее Симпсон отказался от него [Назаров, 91, стр. 14; Симпсон, 48, стр. 156], поэтому использование его в несколько других рамках не приведет к двусмысленностям.
Рассматривая симбиогенную и колониальную теории происхождения эукариот, переход от факультативности к облигатности в симбиозе гриба и водоросли (лишайники), колониальный путь перехода от одноклеточности к многоклеточности у животных и растений, гипотезу автономизации клеточных и генных структур в эволюции вирусов и другие аналогичные примеры, можно прийти к выводу, что мегаэволюционные изменения связаны с изменением степени самостоятельности биологических объектов и, как следствие, с возникновением новых форм и уровней организации живого. При этом очевидно, что в большинстве процессов, являющихся мегаэволюционными механизмами (автономизация, симбиогенез, колониализация), происходит постепенная облигатизация взаимоотношений, сложившихся между исходными бионтами в результате единственного и мгновенного события (например, клеточного каннибализма в случае формирования полового процесса, пожирания предэукариотической клеткой прокариотических эндосимбионтов, отделения участков ДНК при возникновении вирусов и т. п.). Это событие названо здесь мегаэволюционной детонацией и представляет собой элементарный процесс мегаэволюции, подобно тому, как генетическая изоляция (и, как следствие, видообразование) является элементарным процессом мезоэволюции, а точковая мутация - микроэволюции.
Во многих случаях мегаэволюционных преобразований (утрата самостоятельности пластидами или симбионтами-компонентами лишайников) мы можем проследить постадийность этого процесса, в других же (переход к многоклеточности) такое отслеживание будет носить характер умозрительного. Все эти процессы объединены тем, что в них происходит либо разделение одного бионта n-порядка на несколько бионтов (n-1)-порядка (автономизация вирусов), либо, напротив, слияние далеких друг от друга (симбиогенез и межвидовая гибридизация) или однородных (становление полового процесса, колониализация) бионтов n-порядка в бионт (n+1)-порядка; иными словами, в процессе мегаэволюции бионт приобретает уровень (n± 1) (таким образом возникали, по-видимому, разные подцарства растений и животных, многие царства и систематические группы более высоких рангов). Мегаэволюцию, таким образом, можно охарактеризовать как процесс изменения уровня организации бионтов (или степени их энергидности, что практически равнозначно).
Если микроэволюция ограничена допустимыми пределами изменения вида, а мезоэволюция - летальностью большинства преобразований, могущих привести к мезоэволюционным изменениям, то мегаэволюция ограничена, по-видимому, только биологической целесообразностью. Исходя из гипотез ортогенетического толка, можно предположить, что основа для мегаэволюционных преобразований заложена в так называемых “спящих генах”, которых в клетках большинство. Атавизмы, таким образом, можно объяснить как проявление генетической памяти о предковой форме, содержащейся в хромосоме потомка. Этим же (причем не только в мега-, но и в мезоэволюции) объясняется и явление филогенетического ускорения.
Из возможных механизмов мегаэволюции отдельно стоит сказать о межвидовой гибридизации, как естественной, так и искусственной. К мегаэволюционным механизмам это явление причислено потому, что в нем происходит слияние двух бионтов в один, но этот механизм имеет и существенное отличие от других: здесь отсутствует постепенная облигатизация, свойственная, например, симбиогенезу, колониализации или автономизации. Иными словами, межвидовая гибридизация (а также процессы, связанные с изменением числа хромосом и отнесенные в данной работе к мезоэволюционным механизмам) целиком представляет собой акт мегаэволюционной детонации. В настоящее время доказана гибридная природа многих групп животных и растений [Назаров, 91, стр. 4; Стеббинс, Айала, 85], что подчеркивает огромное значение этого механизма.
3. 2. 5. Особенности сингенеза.
Сингенезом называется процесс исторического развития экосистем и всей биосферы. К ведению учения о сингенезе относятся, в первую очередь, периодические смены флор и фаун в процессе исторического развития жизни на Земле. Не вдаваясь в геологические, астрономические и другие подробности, отметим здесь только некоторые закономерности этого процесса. Самым существенным тут является зависимость менее масштабных явлений от более масштабных и явление эволюционного отражения. Исходя из этого тезиса, можно сказать, что три уровня биогенетической эволюции живой природы находятся в зависимости от эволюции природных сообществ, эволюция сообществ - от климатических условий, климат - от глобальных теллурических явлений, а те, в свою очередь, находятся под влиянием космических факторов циклического и ациклического характера. Действительно, модели влияния “космос - абиотические факторы среды - биологические процессы” можно проследить на всех уровнях процессов, связанных с периодическими явлениями космического уровня: суточные изменения, связанные со вращением Земли вокруг своей оси, приливно-отливные процессы, вызываемые лунными циклами, сезонные изменения, основанные на годовом цикле Земли, гелиобиологические явления, вызываемые периодами повышения и понижения солнечной активности и связанные с 11,8-летним юпитерианским годом, и т. п. Причем очевидно, что чем выше уровень космического цикла, тем больше его влияние на абиотические, а, следовательно, и на биотические факторы Земли. Поэтому есть все основания думать, что наиболее глобальные изменения биосферы связаны с наиболее продолжительными космическими циклами, такими, как периоды вертикальных колебаний солнечной системы около галактической плоскости (“галактические сутки” продолжительность около 30 млн. лет), галактический год (180 млн. лет) [Назаров, 91, стр. 128; Красилов, 86] и даже пульсации Вселенной (во всяком случае, скорость разбегания Галактик и степень их удаленности друг от друга). Также в той или иной степени влиять на эволюцию могут периодические явления, связанные с изменением магнитного статуса Земли [Николаев, 91].
Имеет место и обратный процесс, когда меньший по масштабности уровень влияет на больший. Поскольку при каждом переходе на другой уровень масштабность процесса убывает, а обратный процесс возникает в ответ на прямое воздействие, это явление можно назвать эволюционным отражением. В ходе прямого процесса часть энергии, возникающей в ходе эволюции космических объектов, воздействует на геологическо-климатическую эволюцию Земли, та, в свою очередь, влияет на сингенетические процессы, а изменения экосистем вызывает изменение на физиологическом и молекулярно-генетическом уровне, которые и ведут к эволюционным изменениям. Но эволюционные изменения на генетическом уровне, в свою очередь, ведут к изменениям морфологии и экологии, что не может не сказаться на состоянии экосистем, а через них - и биосферы в целом (см. рис. 6); затем происходят дальнейшие затухающие колебания.
Из логики подобного построения следует, что это, в свою очередь, влияет и на астрофизические процессы, однако постепенная потеря энергии от уровня к уровню имеет следствием то, что это влияние заметить весьма проблематично. Тем не менее, антропическое воздействие на Землю может сказаться и на космических явлениях, хотя бы в масштабе Солнечной системы.
Сопоставление палеонтологических и астрономических данных позволило проследить имеющую место корреляцию между периодом вертикальных колебаний солнечной системы (галактическими сутками) и пиками наибольшего вымирания в различных группах живых организмов (30
+4 млн. лет). Что интересно, в палеозое периодичность вымираний была около 34 млн. лет, а в мезозое и кайнозое уменьшилась до 26 млн. лет [Назаров, 91, стр. 130-131], что может свидетельствовать о постепенном сокращении продолжительности “галактических суток” и, возможно, об относительной молодости Галактики, так как после достижения “среднего возраста” колебания, вероятно, должны затухать. На основании этого, а также исходя из того, что скорость мезоэволюции, по-видимому, возрастает, можно сделать вывод (чисто гипотетический), что скорость мезоэволюции каким-либо образом коррелирует с вертикальными колебаниями солнечной системы.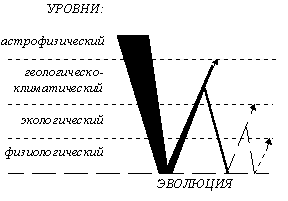
Рис. 6. Эволюционное отражение (толщина стрелки показывает масштабность воздействия).
Возможно, галактический год, как и год земной, характеризуется своеобразными “галактическими сезонами”, которым свойственен больший или меньший уровень радиации и/или какие-либо другие черты, и от этого также зависит эволюционный процесс и другие явления, происходящие на Земле (см. рис. 7). Вероятное с инерционным замедлением скорости вращения галактики может быть связано постепенное замедление мегаэволюционного процесса. Исходя из этих и подобных закономерностей, можно сказать, что сингенетический процесс,
по-видимому, цикличен или, если быть точнее, спирален, то есть последующий виток повторяет предыдущий на более высоком уровне.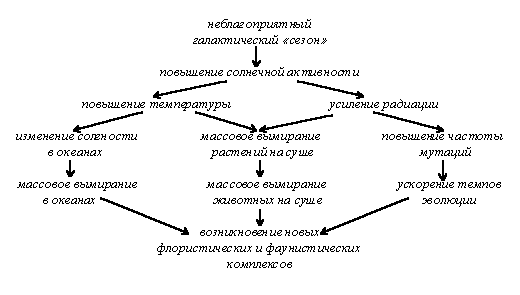
Рис. 7. Возможный механизм периодических вымираний.
Непериодические явления космического уровня, такие, как взрывы сверхновых, ведущие к повышению уровня радиации во много раз [Назаров, 91, стр. 116-118; Красовский, Шкловский, 57, стр. 198], столкновение с Землей астероидов или комет, непосредственно влияющее на биосферу [Назаров, 91, стр. 121-122], и др., тоже, несомненно, внесли (или, по крайней мере, могли внести) свой вклад в катастрофические изменения природных сообществ Земли. Несомненным образом влияя на эволюцию сообществ, катастрофы (ни экстрателлурические, ни теллурические, - хотя последние, вероятно, тоже имеют неземную природу), по-видимому, не способствуют напрямую макро- (мезо- и мега-) эволюции, но позволяют вновь возникающим формам с большей вероятностью выжить и занять свободные экологические ниши. Тем не менее, процессы, связанные с повышением уровня радиации, даже не особо крупными, могут и непосредственно влиять на эволюцию, повышая скорость вымирания отдельных таксонов и увеличивая частоту мутаций, в том числе и макроэволюционного масштаба. Как и для других уровней эволюционного процесса, для сингенеза характерно явление преадаптации: прогрессивные группы возникают в эпохи господства более примитивных, но более приспособленных к данным условиям групп. При изменении условий и вымирании значительной части представителей господствующей группы на первый план выходит эволюционно более продвинутая группа.
Можно отметить еще некоторые особенности сингенеза. Так, несоответствие флористических и фаунистических эр и периодов (флористические изменения обычно предшествуют фаунистическим), а также меньшие масштабы вымирания среди растений, чем среди животных, говорят о том, что растения первыми реагировали на космические изменения, но количество способствующих вымиранию факторов для них было меньше, тогда как на животных влияли также изменения во флоре.
При разговоре о глобальных флористических и фаунистических изменениях невозможно не сказать о значении этого факта в доказательстве ортогенетических концепций. Так, объяснить это явление и прогрессивную эволюцию вообще только естественными (микроэволюционными) факторами невозможно в принципе: в случае, если бы эволюционный процесс направлялся только естественным отбором, эволюция завершилась бы, по крайней мере, на прокариотическом уровне, поскольку известно, что прокариоты обладают всеми мыслимыми способами обмена веществ. Не будь у живой природы имманентного стремления к совершенствованию, все существующие в природе ниши были бы заняты бактериями и археями, что вполне поддерживало бы общебиосферный гомеостаз. Иными словами, уже бактериальная биосфера была вполне самодостаточной и не нуждалась ни в каком дальнейшем совершенствовании путем естественного отбора [Назаров, 91, стр. 235-236]. Более того, в любой момент своего существования компоненты биосферы гармонично связаны друг с другом, и (кроме как в связи с катастрофическими изменениями оболочек Земли) в биологическом прогрессе нет видимой необходимости. Тот факт, что прогрессивная эволюция, тем не менее, идет, неопровержимо свидетельствует, на наш взгляд, о направленности и предопределенности, имеющей место в эволюционном процессе.
3. 2. 6. Эволюция процесса размножения.
Очевидно, что становление того или иного конкретного способа полового размножения (место редукции, соотношение мужских и женских гамет и т.д.) в различных группах происходило самостоятельно. Однако обычно считают, что, во-первых, разделение на два пола если и полифилетично, то происходило относительно немного раз (например, при формировании царств, о чем свидетельствует, в частности, своеобразие полового механизма у инфузорий [Беклемишев, 79] и многих настоящих грибов [Гарибова и др., 75, стр. 82; Жизнь растений; Курс низших растений, 81; Лемеза, Шуханов, 94, стр. 138]), а, во-вторых, что эволюционируют, в основном, особенности полового процесса, а сама его сущность - слияние мужской и женской гаметы - остается неизменной. Тем не менее, ряд фактов (и, прежде всего, различия в способах определения полов даже в пределах одного типа [Айала, Кайгер, 87; Генетика, 91; Медведев, 76]) свидетельствует, что разделение на два пола происходило самостоятельно не только в разных типах одного царства, но и в разных классах (среди позвоночных) и даже отрядах (среди насекомых), а, следовательно, что эволюция крупных таксонов в значительной степени протекает безо всякой зависимости от полового размножения.
Одним из самых любопытных примеров этого плана является отличие генетических характеристик полов в двух ветвях позвоночных - диапсиды и синапсиды (не говоря о частых случаях перехода одного пола в другой у низших позвоночных - рыб и амфибий): так, у синапсид (млекопитающих) гетерозиготны самцы, а у диапсид (птиц, архозавров, лепидозавров) - самки [Айала, Кайгер, 87; Генетика, 91; Медведев, 76], что свидетельствует о том, что разделение на полы происходило независимо друг от друга, по крайней мере, в диапсидной и синапсидной ветвях позвоночных (этот факт является косвенным свидетельством происхождения синапсид и диапсид от разных видов рипидистий). Вообще, если рассматривать группы с различными способами определения пола, можно прийти к выводу, что, допустим, в группе, выделяемой в настоящее время как класс или надкласс Insecta, возникновение полов происходило независимо, по крайней мере, четырежды: у перепончатокрылых (определение пола по признаку гапло- или диплоидии), чешуйчатокрылых (гомозиготность у самцов), двукрылых (по крайней мере, у дрозофилы со сложной системой доминантности женских либо мужских признаков) и других насекомых (гомозиготность у самок) [Генетика, 91; Медведев, 76], - а, скорее всего, гораздо больше раз.
По-видимому, группы с гомозиготностью самцов или самок возникали в результате сложных мутаций половой хромосомы, тогда как группы с гаплодиплоидным типом определения пола или с системой, подобной системе полов у дрозофилы - в результате анеуплоидии (все эти процессы носят мезоэволюционный характер). Однако существуют группы, где формирование полов имеет основу не генетическую, а, например, экологическую [Генетика, 91; Медведев, 76].
Традиционно принято считать, что половое размножение стимулирует эволюцию, так как лишь у организмов с половым размножением действуют такие общеизвестные механизмы эволюции, как дрейф генов и поток генов, а также рекомбинантная изменчивость и кроссинговер [Грант, 80]. Однако можно заметить, что эти явления играют роль только на микроэволюционном уровне. Кроме того, очевидно, что, если разделение на полы возникает, по крайней мере, при эволюции внутри типа, то самые крупные эволюционные изменения происходят без участия полового процесса, причем именно при изучении эволюции классов, типов и выше мы сталкиваемся с наибольшими "белыми пятнами эволюции" и наиболее внезапными преобразованиями [Самовар, 91]. Из этого можно сделать единственный очевидный вывод: половой процесс отнюдь не стимулирует эволюцию, а, напротив, прекращает мега- и мезоэволюционные процессы и подключает микроэволюционные.
Следует рассмотреть наиболее вероятную эволюционную цепочку при становлении полового размножения (см. рис. 8). Разумеется, наиболее примитивной формой размножения следует назвать бесполое, при котором каждая клетка способна дать полноценную особь (вегетативное размножение). Такую форму размножения мы можем наблюдать у одноклеточных и многих колониальных организмов, а из многоклеточных - у губок, кишечнополостных, многих растений, грибов. Следующая ступень эволюции размножения - размножение с дифференцированными группами клеток (клубнями, “усами” и др.) или отдельными клетками бесполого размножения (спорами, или агаметами), которое можно обнаружить в той или иной форме у всех высших растений, а также у фораминифер [Беклемишев, 79, стр. 25] (у животных этот способ размножения не получил распространения). Агаметы, возможно, выделились из плотных групп однородных клеток вегетативного размножения, большинство из которых впоследствии утратили функцию размножения и стало выполнять функции спорангиев. В пользу происхождения гамет от генеративных агамет свидетельствуют также такие явления, как андрогенез (напр., у кукурузы, наездников) и гиногенез (напр., при скрещивании разных видов карасей), а также возможность вызывания у многих видов искусственного партеногенеза путем стимуляции яйцеклетки. Эти факты говорят об изначальной равноценности обоих типов гамет и о первоначальной факультативности оплодотворения.
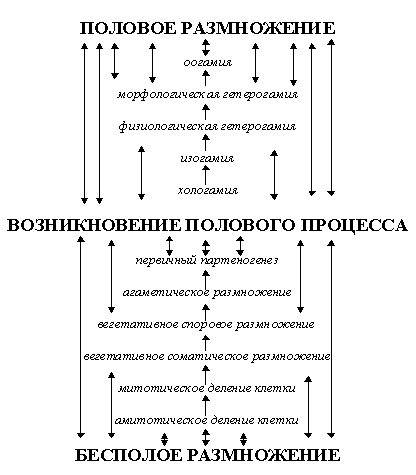
Рис. 8. Эволюция процесса размножения (боковые ветви и совмещение разных способов не учитываются).
Из бесполого размножения с помощью спор напрямую можно вывести первичный партеногенез. Первичность партеногенеза во многих (если не во всех) зарождающихся крупных таксонах решает (вместе с теорией онтомутаций) ранее непреодолимую проблему “многообещающих уродов” (макромутантов, у которых возникают крупные фенотипические изменения, не ведущие к летальности и впоследствии, как предполагалось, становящиеся преадаптациями): как им удается найти брачного партнера со схожим генотипом и при этом избежать близкородственного скрещивания в дальнейшем [Назаров, 91, стр. 143], - так как при партеногенетическом размножении такой проблемы просто не существует. Кроме того, партеногенез, хотя и происходит с помощью гамет, можно рассматривать как видоизмененное споровое, или агаметическое, размножение. Косвенно о преемственности бесполого соматического, бесполого агаметического (спорового) и полового размножения свидетельствует также, например, явление соматического эмбриогенеза у губок.
На каком-либо этапе у различных групп организмов, имеющих гаплоидные агаметы, происходило слияние агамет, приобретающих, таким образом, функции гамет. При этом гаметы сперва были одинаковыми морфологически и физиологически (изогамия, а в случае одноклеточных организмов сперва хологамия), затем приобретали физиологические отличия, при которых сливаться могли только гаметы противоположных знаков, затем одна из гамет становилась больше другой, и, наконец, возникала классическая оогамия [Петров, 74]. Не исключено, что возникновение полового процесса было связано с адаптацией к клеточному каннибализму, что напоминает мегаэволюционные механизмы (возникновение диплоидной формы жизни из двух гаплоидных). В таком случае, акт каннибализма соотносим с мегаэволюционной детонацией, а после нее следует процесс постепенной облигатизации оплодотворения. Оогамия, судя по всему, возникла в результате мезоэволюции, тогда как гетерогаметизация - процесс явно микроэволюционный.
Разумеется, обнаружить полную цепочку эволюции процесса размножения в развитии какого-либо организма практически невозможно, так как отдельные стадии зачастую выпадают. К тому же, у огромного количества групп имеет место сохранение примитивных форм размножения при наличии прогрессивных. Так, например, кишечнополостные, губки, высшие растения, приобретя способность к половому размножению, весьма часто "пользуются" и вегетативным размножением, многие растения чередуют стадии с половым и споровым размножением, а у такой прогрессивной группы, как млекопитающие, бесполое размножение сохранилось в видоизмененной форме, в виде деления зиготы с возникновением однояйцевых близнецов (у броненосцев, например, эта форма размножения достаточно распространена).
3. 2. 7. Взаимовлияние эволюционных механизмов.
Хотя мега-, мезо- и микроэволюционные механизмы действуют самостоятельно, они взаимно влияют друг на друга. Так, на определенном этапе к мегаэволюционным механизмам подключаются мезоэволюционные, а к мезоэволюционным - микроэволюционные. Наиболее важным здесь является уже упоминаемое ранее явление неотении, поскольку, как отмечалось ранее, высокая специализация затрудняет эволюцию, а явление неотении позволяет уменьшить специализацию и, таким образом, усилить действие микроэволюции. Неотения, возникшая у партеногенетически размножающихся организмов (педогенез), является, по всей видимости, базовой на момент возникновения полового процесса [Захваткин, 49]. Кроме того, механизмы всех уровней эволюции оказывают влияние на виды одновременно, хотя и каждый по-своему, и, тем не менее, достаточно просто путем логических умозаключений прийти к выводу, каким путем произошло то или иное преобразование. От этого зависит и систематическая ценность различных признаков.
Можно отметить следующие типы взаимовлияния различных уровней эволюционного процесса. Во-первых, мега- и мезоэволюционные преобразования подвергаются впоследствии микроэволюционному отбору. Во-вторых, завершение действия мегаэволюционных механизмов приводит к подключению мезоэволюционных, а завершение действия мезоэволюционных - к подключению микроэволюционных. В-третьих, как уже отмечалось, некоторые мезоэволюционные преобразования сами по себе могут способствовать усилению действия микроэволюции. В-пятых, одно и то же генетическое изменение может привести к изменениям на разных уровнях. В-шестых, мегаэволюционные преобразования (как, например, приобретение предэукариотами бактериальных эндосимбионтов) могут являться преадаптациями для мезоэволюционных преобразований (в приведенном примере - связанных со становлением митоза и других). Наконец, могут быть и иные способы взаимовлияния.
Можно заметить и некоторую постепенность в переходе от микроэволюционных механизмов к мегаэволюционным через мезоэволюционные. С одной стороны, трудно провести четкую границу между микро- и макромутациями, различие между которыми является одним из разграничивающих моментов между микро- и мезоэволюцией (в ряде случаев это различие количественное, а не качественное), а даже равнозначные микромутации структурных и регуляторных генов ведут к совершенно разным последствиям. Далее, транспозоновый перенос, ведя, как и мутации, к моментальным перестройкам в геноме, уже имеет некоторые мегаэволюционные черты, так как связан с информационным взаимодействием разных (в том числе далеких друг от друга) видов. Гибридизация, представляя собой объединение двух бионтов в один, не характеризуется постепенной облигатизацией взаимоотношений. Наконец, симбиогенез в полной мере обладает мегаэволюционными чертами. Аналогичные сопоставления можно сделать и по другим механизмам (колониализация, автономизация и т. д.). Это говорит, однако, не о единстве и взаимной сводимости разных уровней эволюционного процесса, а, напротив, о еще большей его дробности, что дает основание подразделять эволюцию на еще большее число уровней, не связанных друг с другом.
Выявление того, с помощью какого механизма возникала та или иная группа живых организмов, позволит унифицировать систематику, так как этот признак является качественным, а не количественным (то есть показывает реальную природу данной группы, а не степень ее отличия от ближайших к ней родственных групп или от предковых форм), и, к тому же, свойственен всем без исключения живым организмам. Возможно, использование такого подхода поможет созданию универсальной системы живых организмов наподобие периодической системы химических элементов, созданной Менделеевым [Московский, 99].
3. 2. 8. Критика концепции искусственного отбора.
Отдельно следует сказать о часто рассматриваемом разграничении эволюционного процесса на эволюцию, связанную с естественным отбором (эволюцию биологических видов), и эволюцию, связанную с искусственным отбором (эволюцию сортов и пород). Согласно таким концепциям, человек противопоставляется всей остальной природе (следствие антропоцентрического мировоззрения, недопустимого для настоящего исследователя природы!), тогда как в действительности человек выступает в роли внешнего (по отношению к популяции, в которой производится отбор) фактора, наряду с другими факторами природы. Противопоставление антропогенного (антропического) фактора биотическому в корне неверно с точки зрения естественного классифицирования: хотя он и весьма значителен в планетарном масштабе, но, подходя с качественной, а не с количественной позиции, неотличим от, например, воздействия муравьев на популяции тлей или от мелиоративной деятельности бобров.
С точки зрения сторонников концепции искусственного отбора, сорта растений и породы животных не подвержены борьбе за существование. Однако в действительности можно сказать, что для них борьба за существование просто проявляется в других формах, нежели для их “диких” родственников, - а именно, в соответствии определенных признаков запросам биологического вида животных Homo sapiens, одного из многочисленных компонентов живой природы.
Таким образом, искусственный отбор представляет собой всего лишь форму естественного, поскольку антропический фактор отличен от биотического только в масштабах и формах воздействия на природу, но не в качественных признаках: и тот, и другой представляют собой изменение окружающего мира живыми организмами (включая человека) с целью приспособления к условиям среды и с помощью своей активной деятельности. Тем более, разделение естественного и искусственного отбора теряет свою актуальность, если признать, что отбор лежит только в основе микроэволюции, так как в этом случае исчезает такое важное отличие, как разные результаты действия двух видов отбора.
1. Процесс исторического развития жизни на Земле можно разделить по уровням организации жизни на онтогенез (индивидуальное развитие отдельных организмов), филогенез (историческое развитие систематических групп живых организмов) и сингенез (эволюция экосистем и биосферы); в филогенезе, в свою очередь, можно выделить три уровня: микроэволюция, мезоэволюция и мегаэволюция.
2. Микроэволюционный процесс представлен постепенными ненаправленными изменениями, основанными на микромутациях и движущих силах микроэволюции, мезоэволюционный - мгновенными перестройками генома, ведущими к генетической изоляции, мегаэволюционный - постепенной облигатизацией нового уровня энергидности, инициированной актом мегаэволюционной детонации.
3. Высший уровень эволюции не сводим к низшему, однако разные уровни влияют друг на друга.
4. Элементарным процессом микроэволюции являются микромутации структурных генов и рекомбинации, движущими силами - поток генов, дрейф генов, борьба за существование и естественный отбор.
5. Микроэволюция имеет обратимый характер и (за редким исключением) не ведет к генетической изоляции и, стало быть, к образованию новых видов.
6. Элементарным процессом мезоэволюции является мгновенная генетическая изоляция, связанная с хромосомными мутациями, мутациями регуляторных генов и горизонтальным переносом генетической информации.
7. Значительную роль в мезоэволюционных преобразованиях играют различные гетерохронии, онтомутации и другие явления, связанные с изменением протекания онтогенетических процессов.
8. Эволюция на макро- (мезо- и мега-) -эволюционных уровнях в значительной степени имеет пре-, а не постадаптативный характер.
9. Элементарным процессом мегаэволюции является мегаэволюционная детонация - одномоментный акт изменения энергидности и уровня организации эволюционирующего бионта, - ведущая затем к постепенной облигатизации сложившейся формы жизни.
10. К мегаэволюционным изменениям можно отнести такие явления, как межвидовая гибридизация, полиплоидизация, колониализация, симбиогенез, а также автоматизация клеточных структур при возникновении вирусов, однако каждый из этих процессов имеет и собственную специфику.
11. По-видимому, магистральные пути мегаэволюции ортогенетически предопределены.
12. В сингенетических процессах менее масштабные явления зависят от более масштабных, следовательно, глобальные смены флор и фаун предопределены циклическими и нециклическими космическими процессами.
13. Имеется и обратное воздействие мене масштабных исторических процессов на более масштабные, но оно ослабляется при переходе на каждый последующий уровень (явление эволюционного зеркала).
14. Наиболее крупными космическими явлениями, влияющими на эволюционный процесс, являются вертикальные колебания солнечной системы относительно плоскости галактики (галактические сутки, длящиеся около 30 млн. лет), вращение солнечной системы вокруг центра галактики (галактический год, длящийся около 180 млн. лет) и процесс разбегания галактик относительно центра сингулярности, причем четко прослеживается корреляция между этими процессами (по крайней мере, первыми двумя) и сменами флор и фаун.
15. Из непериодических явлений существенное влияние на земную биосферу оказывают взрывы сверхновых, а также столкновения с крупными небесными телами наподобие комет или астероидов.
16. Для сингенеза характерно явление преадаптации, проявляющееся в том, что прогрессивные группы возникают задолго до окончания периода господства более примитивных.
17. Флористические изменения происходят раньше фаунистических, что можно объяснить тем, что растения более чувствительны к космическим изменениям, чем животные (так как их жизнедеятельность напрямую зависит от уровня освещенности и других факторов, берущих начало в космосе), тогда как большие масштабы вымирания среди животных, чем среди растений, вызваны зависимостью животных не только от абиотических факторов среды, но и от состояния растительности.
18. Прогрессивная эволюция является свидетельством ее ортогенетического характера.
19. Половой процесс как процесс слияния гамет широко полифиличен, что доказывается, прежде всего, множественностью способов определения пола в разных группах живых организмов.
20. Возникновение полового процесса связано с мегаэволюционной детонацией в виде слияния агаметических клеток бесполого размножения (“спор”) и постепенной облигатизацией такого способа взаимодействия; таким образом, партеногенез можно рассматривать как атавизм, восходящий к более древней форме размножения.
21. Неотения, возникшая у партеногенетически размножающихся организмов (педогенез), является, по всей видимости, базовой на момент возникновения полового процесса.
22. Уровни эволюции не вытекают один из другого, но сложно и многосторонне влияют друг на друга.
23. Механизмы эволюции, сочетающие черты разных уровней, не являются переходными формами, но свидетельствуют о том, что даже разделение на три уровня еще не дает полного представления о многообразии путей эволюции.
24. Представления об искусственном отборе как особой форме эволюционного механизма вызвано антропоцентрическим мировоззрением, а не естественными отличиями этого процесса от естественного отбора.
§ 3. 3. УМОЗРИТЕЛЬНЫЙ АНАЛИЗ ЭВОЛЮЦИОННОЙ СИСТЕМАТИКИ:
ВОЗМОЖНЫЕ ПУТИ ЭВОЛЮЦИИ ЖИЗНИ.
3. 3. 1. Происхождение прогенота.
Жизнь можно определить как способ взаимодействия материи, энергии и информации, характеризующийся способностью системы к самовоспроизведению по единой информационной матрице (информационный критерий), обмену веществом, энергией и информацией с окружающей средой (экологический критерий), а также индивидуальному и историческому развитию вследствие случайных или закономерных изменений в информационной структуре (эволюционный критерий). Информационный критерий имеет следствием рождение, умирание и наследственность, экологический - дыхание, питание, выделение, рост и раздражимость, эволюционный - изменчивость, онтогенез и филогенез. На наш взгляд, эти три критерия являются необходимыми и достаточными для понимания сущности жизни и введение четвертого, химического критерия (белковая, или нуклеопротеиновая природа) только неоправданно сужает поле биологических исследований, не давая потенциальной возможности для изучения небелковых форм жизни, если таковые будут обнаружены. Напротив, ликвидация любого из трех вышеперечисленных критериев ведет к неоправданному расширению поля исследований и включению в него явно лишних объектов. Так, без информационного критерия живыми существами можно было бы назвать биогеоценозы, планеты, звезды и т. д., что хотя и может оказаться логичным с мистической и философской точки зрения, но не оправдано биологически; без экологического критерия к живым существам можно было бы отнести большинство информационных объектов (человеческие мысли, компьютерные базы данных и т. п.), что тоже согласуется с некоторыми мистическими представлениями (теория “Мимов”), но не с биологией; наконец, без эволюционного критерия живыми существами являлись бы машины, сконструированные для изготовления себе подобных, что пока что является объектом хотя и фантастики, но научной. Приведенное же выше определение жизни позволяет по-новому взглянуть на проблему происхождения жизни на Земле, до сих пор являющуюся одним из важнейших дискуссионных вопросов теории эволюции.
В настоящее время наибольшей популярностью пользуются гипотезы химического (автохтонного) и космического (панспермия) появления жизни на Земле (именно на Земле, так как космическая теория не снимает вопроса о происхождении жизни вообще) [Васильев, 65; Войнович, 88], причем традиционные биохимические теории (прежде всего, Опарина [Опарин, 57, 66]) сталкиваются со все большими затруднениями, тогда как космические теории получают все новые подтверждения. Так, уже доказана возможность переноса сложных органических соединений через космическое пространство на метеорных телах, а также возможности ряда микроорганизмов выдерживать условия открытого космоса в анабиотическом состоянии [Все живое на Земле произошло из нефти! 99]. Сравнительно недавно получила подтверждение гипотеза, согласно которой жизнь может существовать в недрах практически любой планеты, так как, во-первых, по мнению американского астробиолога Д. Ноэвера и английского генетика С. Джонеса, земные условия можно считать уникальными только в том случае, если рассматривать поверхность, а не недра, во-вторых, в нефти обнаружены такие своеобразные по своей биохимии организмы, как археи, а, в-третьих, новые данные позволяют предположить, что нефть образует не отдельные скопления в толщах литосферы, а особый слой в строении Земли (и, возможно, других планет), причем, по мнению американских астрономов Дж. Клампета и Т. Голда, нефть отнюдь не является продуктом жизнедеятельности растений, как это предполагалось ранее [Все живое на Земле произошло из нефти! 99]. В таком случае, жизнь (или, во всяком случае, предбиологические системы типа нуклеиновых кислот) может оказаться почти таким же древним явлением, как и Вселенная, и при объяснении ее происхождения в масштабах Космоса просто невозможно не предположить влияние некоего "Высшего Разума", будь то какое-то сверхъестественное существо или закон природы, позволивший возникнуть первичной форме жизни с универсальными свойствами, такими, как, например, генетический код и способность к эволюционированию.
Как уже было сказано, современные теории химического происхождения жизни на Земле страдает рядом внутренних противоречий, которые свидетельствуют о невозможности такого пути возникновения живых организмов. Так, можно отметить, что "опаринским" путем сложные органические соединения могли возникнуть только в восстановительной среде, а сохраниться - только в окислительной, что создает главный парадокс. Второй парадокс, подобный вопросу "о курице и яйце", заключается в том, что синтез нуклеиновых кислот невозможен без белков, тогда как белки кодируются нуклеиновыми кислотами. Парадоксальным является также, например, построение всех белков только из двадцати аминокислот из более сотни известных, причем из их левоасимметричных форм (вариант решения этой и некоторых других проблем указан в дополнении к данному разделу) , и многие другие моменты. Кроме того, вода является сильным растворителем, что тоже говорит не в пользу "гипотезы первичного бульона" (во всяком случае, не в ее традиционном представлении) [Жизнь - как она возникла? 85, стр. 40-45]. Все это, очевидно, свидетельствует в пользу того, что в условиях земной поверхности (в том числе океана) жизнь (во всяком случае, синтез белка по нуклеиновым матрицам) возникнуть не могла.
Наша гипотеза о происхождении жизни на Земле представляет собою синтез космической и химической теорий с теорией последовательных эндосимбиозов, примененной не только к формированию эукариотического типа организации клетки, но и к возникновению прогенота. Поскольку, как уже говорилось, самопроизвольное появление синтеза белков и нуклеиновых кислот на Земле вряд ли было возможно, остается предположить, что он возник в условиях, отличных от земных, и способами, пока что не выясненными окончательно. Так называемые “споры жизни”, фигурирующие в теории панспермии (назовем их витоспорами), представляли собой, по-видимому, систему, включающую нуклеиновую кислоту (ДНК либо, вероятнее, РНК) и белковую или липопротеиновую оболочку [Маргелис, 83, стр. 19] и способную при попадании в подходящие условия начать процесс самокопирования, то есть наделенную информационным признаком жизни; таким образом, витоспоры могли напоминать вирион.
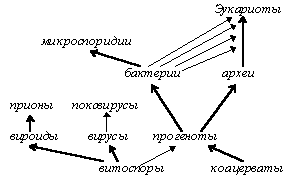
Рис. 9. Возможная схема возникновения основных форм жизни
(приобретение эндосимбионтов обозначено треугольными стрелками).
К определенному моменту (около 3,5 - 4 млрд. лет назад) химическая эволюция на Земле привела к возникновению коацерватоподобных образований, способных к обмену веществ и росту, то есть имеющих экологический признак жизни, но неспособных к копированию входящих в их состав молекул [Опарин, 57, 66]. В результате бомбардировки земной поверхности ДНК- и/или РНК-содержащими витоспорами различные разновидности последних (или же витоспоры единственного типа, впоследствии эволюционирующие внутри клетки) неоднократно вступали в эндосимбиозы с присутствующими на Земле коацерватами, что позволило совместить информационный и экологический критерии живой материи и привело к появлению третьего компонента - способности к эволюционированию. Вероятно, предбиологическая эволюция органического вещества на Земле и формирование симбиоза витоспор и коацерватов во многом была связана с вращением галактики, в ходе которого периодически менялся уровень космической радиации, влияющей на скорость эволюционного процесса [Шкловский, 73; Назаров, 91, стр. 117]. С вращением же галактики, а так же с еще более глобальным процессом разбегания галактик относительно центра сингулярности, связано, по-видимому, и периодическое попадание различных планет в потоки вещества, несущего витоспоры.
Механизм первоначальной симбиотической деятельности коацервата и витоспор неясен, однако очевидно, что ДНК-витоспоры дали начало нуклеоиду прогенота (и через него - геному бактерий, архей, эукариот и их прокариотических эндосимбионтов), а РНК-витоспоры (возможно, разные их формы) - матричным, транспортным и рибосомным РНК клеток (вероятно, первичными были структуры, напоминающие тРНК) и всему рибосомному комплексу, отличающему клеточные формы жизни от неклеточных (вирусов) (процесс симбиогенного возникновения прогенота и эукариот, а также возникновение основных форм жизни - см. рис. 9).
Впрочем, не исключена и несколько иная картина: витоспоры могли содержать только один тип нуклеиновой кислоты (одноцепочечная или двухцепочечная ДНК или РНК), а другие типы могли возникнуть уже в результате земной эволюции витоспоры (самостоятельной или, скорее, в комплексе коацерват-витоспора). Эта версия косвенно подтверждается явлением прямой и обратной транскрипции [Вирусология, 89; Жданов, 66], а также существованием сложных поксвирусов - вероятных потомков коацерватов с одним типом нуклеиновой кислоты, - самостоятельная же эволюция витоспор кажется нам маловероятной.С момента объединения витоспор и коацерватов в единый симбиотический комплекс началось развертывание на Земле предопределенной природой эволюционной программы, подобной космической эволюции химических элементов как заполнению вакансий периодической системы от водорода к тяжелым элементам. Говоря о предопределенности эволюции, мы не имеем в виду, что изначально было определено все нынешнее многообразие видов (эта гипотеза полностью отвергала бы действие окружающей среды и случайных факторов и означала бы, что на всех планетах, имеющих биосферу, проживают генетически идентичные виды, что противоречило бы теории вероятности): мы считаем лишь, что еще не раскрытые человеком законы природы направляют эволюцию в любых условиях по одним и тем же магистральным путям, подобно тому, как периодическая система отнюдь не находится в противоречии с фактом существования изотопов. Однако точно убедиться в правильности того или иного взгляда на эволюцию в космическом масштабе мы сможем только в том случае, если обнаружим более или менее высокоорганизованную нуклеопротеиновую (или иную) жизнь на других планетах.
Выше говорилось, что синтез белков по матрице нуклеиновых кислот вряд ли мог возникнуть в земных условиях. В последнее время мы заинтересовались этапами ранней эволюции жизни, связанными с такими глобальными явлениями, как возникновение разных типов аминокислот (в частности, их левосторонней асимметрии) и нуклеотидов, а также генетического кода. Не вдаваясь в подробности хода наших мыслей (этому, вероятно, будут посвящены наши дальнейшие работы), приведем здесь некоторые сделанные нами выводы по этому вопросу, хотя они и требуют еще дальнейшей доработки (особенно по биохимическому объяснению нижеперечисленных событий).
1. Левосторонняя асимметрия аминокислот, во многом создавшая условия для появления белковой жизни, может объясняться единственным способом, не вступающим в противоречие с теорией вероятности: все левосторонние “волшебные аминокислоты”, входящие в состав белков, происходят от единственной левоасимметричной “волшебной аминокислоты” простого строения в результате ряда своеобразных “химических мутаций”.
2. Первичная нуклеиновая кислота состояла только из одного типа нуклеотидных мономеров, кодирующих единственную аминокислоту (по-видимому, это были нуклеотидные остатки цитозина), и имела (в том случае, если первичная нуклеиновая кислота была двухцепочечной) либо синтезировала (если она была одноцепочечной) негативную (не являющуюся матрицей) цепочку комплиментарных нуклеотидных остатков.
3. Комплексы нуклеиновой кислоты и аминокислоты напоминали, вероятно, современные транспортные РНК.
4. Параллельно идущие “химические мутации” нуклеотидов и аминокислот постепенно привели к возникновению современного их числа.
5. Наиболее простая мутация процесса матричного синтеза привела к считыванию информации с негативной нити РНК или ДНК, либо связанной, либо не связанной с позитивной пространственно.
6. Пять главных (а также аномальные) нуклеотидов сформировались раньше, чем двадцать “волшебных аминокислот).
7. На ранних этапах эволюции жизни аминокислоты кодировались двумя, а не тремя, нуклеотидами. Третий нуклеотид триплета представлял собой аналог современных генетических “знаков препинания”, но не между генами, а между кодонами. В это время белки состояли только из 7-9 типов аминокислот (тогда как максимальное количество аминокислот, которые можно закодировать дуплетным кодом, равно шестнадцати).
Эти и другие выводы можно сделать, основываясь на сопоставлении структуры триплетов современного генетического кода, однако для более полной научной проработки данного вопроса нужно более подробно ознакомиться со структурами и свойствами нуклеиновых кислот и аминокислот. Решение этой проблемы позволит описать возникновение нуклеопротеиновой жизни как таковой, а не только в масштабах земной биосферы.
. Происхождение и эволюция вирусов.Некоторые из витоспор или их симбионтных потомков могли неоднократно переходить к генетическому паразитизму, встраиваясь в геном прогенота и реплицируясь за счет генов и рибосомных механизмов хозяина. Постепенно упрощаясь (или, во всяком случае, не прогрессируя) под влиянием паразитизма и эволюционируя (в основном, идиоадаптивно) параллельно с хозяевами, а также передаваясь от одного вида к другому и участвуя в процессе горизонтального переноса, они могли дать начало значительному числу современных ДНК- и РНК-содержащих вирусов. Другие вирусы могли возникнуть в результате автономизации и редукции субклеточных и генных структур, которые, таким образом (если принять симбиогенную теорию во всей ее полноте, включая то, что описано в прошлом разделе), тоже ведут свой род от симбионтных витоспор. При этом следует отметить, что и первым, и вторым путем возникновение различных вирусов, очевидно, происходило неоднократно, что дает основание считать, что вирусы - широко полифилетичная группа. Исходя из этого, в классификации вирусов и вирусоподобных организмов основным критерием следует, по-видимому, считать тип генома и способ его реализации (стратегии реализации генетической информации в основных группах вирусов можно найти в приложении 1 или в первоисточнике [Жданов, 66, стр. 49 - 52]).
Отдельно следует сказать о таких группах вирусов, как прионы и вироиды, с одной стороны, и поксвирусы - с другой. Прионы, по-видимому, полностью лишены нуклеиновой кислоты [Жданов, 66; Вирусология
, 89], поэтому долгое время механизм их репликации оставался не выясненным. Для объяснения этого феномена предлагалась даже такая версия, как обратная трансляция нуклеиновой кислоты по белковой матрице, что вряд ли возможно с биохимической точки зрения. Сейчас считается наиболее вероятным, что прионный белок инициирует действие каких-либо генов клетки-хозяина, на матрице которых записан код прионного белка [Жданов, 66; Вирусология, 89]. Возможно, это говорит о том, что прионы возникли из первичного белка, входившего в состав витоспоры, нуклеоид которой встроился позднее в клеточный геном в качестве “спящего гена”, активирующегося только при контакте с соответствующим белком. Аналогичную природу - видимо, переходную между витоспорой и прионом - имеют и вироиды: они содержат короткую цепочку РНК, но процесс репликации у них, как и у прионов, связан, по-видимому, не с РНК, а с белком [Жданов, 66; Вирусология, 89]. На наш взгляд, эти факты свидетельствуют о том, что между вирусным белком и вирусной нуклеиновой кислотой существует сложная и неоднозначная связь, которая может объясняться единством их возникновения в сильно специфических (возможно, внеземных) условиях.Напротив, поксвирусы (вирусы оспы) характеризуются большой величиной генома, сопоставимой с величиной генома риккетсий, хламидий и микоплазм, автономностью репликации и транскрипции, отсутствием сплайсинга, сложностью строения и химического состава [Жданов, 66; Вирусология, 89]. От мелких прокариот они отличаются только отсутствием рибосомного аппарата и, стало быть, зависимостью трансляции белка от рибосомного аппарата хозяина. Как уже упоминалось, поксвирусы могут быть потомками коацерватов, вступивших в симбиоз только с ДНК-витоспорой. Однако они могут также являться упрощенными в связи с паразитическим образом жизни прокариотами типа риккетсий или хламидий, либо прокариотическими же эндосимбионтами эукариот наподобие пластид или митохондрий с редуцированным вследствие паразитизма рибосомным аппаратом.
При рассмотрении происхождения различных групп вирусов логично использовать не термины “моно- и полифилия”, а аналогичные им термины “гомо- и гетерофилия”, так как в ряде случаев при фактической полифилии какой-либо группы вирусов возникали все ее представители из схожих структур и одинаковым образом (гомофилия). Возникновение из разных структур, но аналогичным образом, равно как и из одинаковых структур, но разными способами, можно называть псевдогомофилией, а совершенно несхожие по путям происхождения вирусы гетерофилетичны.
Вирусы представляют собой настолько уникальные объекты, что у многих исследователей даже возникает вопрос, правомерно ли считать их живыми существами, так как к ним слабо применимы многие общебиологические понятия, такие как вид, популяция и т. п. [Жданов, 66, стр. 7-9]. Тем не менее, вирусы способны к размножению и эволюции, а в комплексе вирус-клетка - и к обмену веществом, энергией и информацией со средой, в том числе с клеткой-хозяином, то есть обладает всеми основными свойствами жизни. Отсутствие же собственных систем синтеза белка [Жданов, 66, стр. 6] является не отличием живого от неживого, а отличием неклеточной формы организации жизни от клеточной. Тем более, если представленная в данной работе космическо-теллурическая гипотеза возникновения жизни верна, вирусы (или, точнее, вирусоподобные существа) можно считать первыми по-настоящему живыми существами на Земле: эволюционно они неразрывно связаны с другими живыми существами и потому не могут рассматриваться иначе, чем в единой системе с ними.
3. 3. 3. Происхождение и ранняя эволюция эукариот.
На ранних стадиях своей эволюции прогенот дал начало двум биохимически и генетически разным группам прокариотических организмов: археям, или архебактериям (эфиры глицерина и фитанола в составе липидов мембраны, кислые полисахариды, белки или псевдомуреин в составе клеточной стенки, РНК-полимераза из 9-12 субъединиц и ряд других особенностей), и собственно бактериям, или эубактериям (в составе клеточной стенки - муреин, РНК-полимераза из 4-8 субъединиц). Вероятно, основная эволюция бактерий протекала в водной среде, а архей - в нефтяной [Все живое на Земле произошло из нефти! 99], хотя некоторые из архей являются водными организмами. Прокариотические организмы (особенно бактерии) дали большое разнообразие форм, имеющих все возможные типы обмена веществ и питания, кроме голозойного (фагогетеротрофного). Многие из бактерий, характеризующихся разными цитологическими и биохимическими особенностями, в разные моменты эволюционной истории вступали в симбиоз с более крупными археями, лишенными плотных клеточных стенок, что в конечном итоге привело к возникновению клеток эукариотического типа организации.
Таким образом, в целом мы придерживаемся, как и в случае с прогенотом, теории о происхождении эукариот в результате последовательных симбиозов, в результате которых клетка-хозяин (этот термин, обычно использующийся при описании внутриклеточного паразитизма, мы будем применять к более крупному компоненту при клеточном симбиозе, так как, во-первых, паразитизм есть тоже форма симбиоза в широком смысле слова, а, во-вторых, этот компонент впоследствии стал основой эукариотической клетки) приобретала митохондрии (в качестве бактерий, родственных митохондриям эукариот, иногда называют паракокков и бделловибрионов), пластиды, а также, возможно, ядра и полярные структуры с наборами микротрубочек [Маргелис, 83, стр. 14]. На основании сходства АТФазы архей и дрожжевых вакуолей и ее отличия от АТФазы других дрожжевых субклеточных органелл, а также в результате сравнения некоторых других ферментов, высказываются даже предположения о симбиогенном происхождении вакуолей и других клеточных структур [Кулаев, 98], причем в симбиоз при сборке эукариотической клетки могли вступать не только археи и бактерии, но и, как и при сборке прогенота, витоспоры и, возможно, какие-либо предбиологические системы, возникшие на Земле химическим путем. Однако главным в возникновении эукариотической клетки событием следует считать именно приобретение микротубулярных структур, обеспечивающих разнообразные формы клеточного движения и ставших преадаптацией для многих важных явлений в жизни эукариот (см. рис. 10), вне зависимости от того, каким путем клетка-хозяин эти структуры приобрела.
По-видимому, клеткой-хозяином являлся какой-то вид архей наподобие Thermoplasma [Маргелис, 83] или, скорее, Halobacterium, что подтверждено сходством 5SРНК эукариот и архей этого рода [Жданов, 66, стр. 39]. Вообще, биохимические отличия архей от бактерий друг от друга столь велики, что по сравнению с ними отличия, например, покрытосеменных растений от млекопитающих ничтожны [Маргелис, 83; Мирабдуллаев], тогда как археи и эукариоты имеют схожие многократно повторяющиеся нуклеотидные последовательности и интроны в геноме, что дает основания считать, что ранее объединяемые в одну империю Prokaryota археи (архебактерии) и бактерии (эубактерии) отстоят друг от друга значительно дальше, чем археи и эукариоты, и может свидетельствовать о противопоставлении архей и эукариот бактериям (иными словами, прокариоты - парафилетическая группа, не отражающая реального родства, а указывающая на уровень организации).
На определенном этапе эволюции предэукариотическая клетка приобрела микротубулярные структуры либо эндо-, либо экзогенно, путем симбиоза с организмами, обладающими такими структурами (Маргелис называет в качестве таких организмов спирохет или спироплазм, причем у спироплазм обнаружен фибриллярный белок с молекулярной массой более 5000, однако его гомология с тубулином пока что не установлена) [Маргелис, 83, стр. 16]. Нам представляется более вероятной гипотеза о симбиогенном приобретении микротрубочек (в таком случае, по крайней мере, один ундулиподиальный экзосимбионт появился у общего предка всех эукариот, а позднее в разных ветвях снова могли происходить подобные же экзосимбиозы; это объяснило бы гетероконтное жгутикование пединеллид и некоторые другие моменты), но говорить с уверенностью об этом у нас пока что нет оснований. В результате приобретения микротрубочек клетки обрели способность к локомоции (исходя из принципов вероятности и экономичности развития, скорее всего, первичными были обособленные от клеток жгутики, которые, постепенно интегрируясь в клетку, дали возможность для псевдоподиальной локомоции, так как более вероятно полифилетическое возникновение просто устроенных псевдоподий, чем таких сложных структур, как ундулиподии - жгутики и реснички; этот момент, пожалуй, можно трактовать как свидетельство экзогенного появления микротрубочек), движению цитоплазмы и фагоцитозу. Фагоцитоз, в свою очередь, дал возможность для приобретения эндосимбиотических партнеров в виде эубактерий, ставших впоследствии митохондриями и пластидами (а также, возможно, ядрами), так как эубактериальные эндосимбионты имели метаболизм, сильно отличающийся от метаболизма архей, и потому, вероятно, не переваривались ими (партнер, принесший, в случае верности симбионтной теории происхождения полярных структур и жгутиков, в клетку предэукариот микротрубочки был, видимо, экзосимбионтом, лишь впоследствии внедрившимся в клетку). Кроме того, приобретение микротрубочек стало преадаптацией к формированию таких важных свойств эукариот, как митоз, половой процесс и мейоз, причем микротубулярные структуры в виде разнообразных клеточных центров стали гарантом правильности расхождения хромосом, организуя нити веретена деления.
Происхождение обособленного ядра может быть связано с поэтапной мембранизацией, однако тот факт, что ядро, как и митохондрии и пластиды, имеет двойную мембрану, позволяет поставить это под сомнение. Не исключено, что ядро приобретено также эндосимбионтно, а собственная ДНК цитоплазмы (то есть клетки-хозяина) была утрачена [Маргелис, 83, стр. 163-164]. Впрочем, организм, считающийся наиболее примитивной современной эукариотой - Pelomyxa [Маргелис, 83; Жданов, 66], лишенная митоза и митохондрий и содержащая прокариотического симбионта, выполняющего их функции, - имеет ядро, но лишен 9+Х-фибриллярных структур, что может трактоваться неоднозначно: либо эндосимбионт, давший начало ядру, приобретался неоднократно, либо ядро все же возникло эндогенно и раньше, чем были приобретены микротрубочки, либо (и эта версия кажется нам в свете нашей гипотезы наиболее убедительной, так как Pelomyxa обладает клеточной подвижностью и, стало быть, по-видимому, имеет белок, гомологичный тубулину, хотя и не структурированный) предок этого амебоидного организма имел жгутик, который, как и в случае со многими другими организмами, не имеющими жгутиковой стадии, интегрировался в клетку и деструктурировался, дав возможность для фагоцитоза, псевдоподиальной локомоции и движения цитоплазмы, но не сформировав митотический механизм.
Важным моментом для понимания происхождения эукариот является также происхождение свойственного им типа генома, который обычно представлен несколькими некольцевыми хромосомами, наделенными гистоновыми белками [Эволюция генома, 86]. Поскольку единственными известными отклонениями от этого типа генома среди эукариот являются динофлагелляты (их называют также мезокариотами за свойственный им тип генома), находящиеся в основании эволюционного ствола трубчатокристовых и имеющие, как и бактерии, кольцевые хромосомы, лишенные или почти лишенные гистонов [Пахомова, 72; Седова], то следует предположить, что множественность хромосом у эукариот возникла в результате полиплоидии (либо, как и половой процесс, в результате клеточного каннибализма), и, таким образом, практически все хромосомы по своему происхождению являются гомологичными. Однако впоследствии эти хромосомы многократно мутировали в независимых друг от друга направлениях, что и привело к нынешнему многообразию форм хромосом и, следовательно, видов эукариот, так как с увеличением генома увеличились и возможности изменчивости (мутации, даже весьма значительные, но происходящие на базе дуплицированных генов и хромосом, могли не вести к летальности).
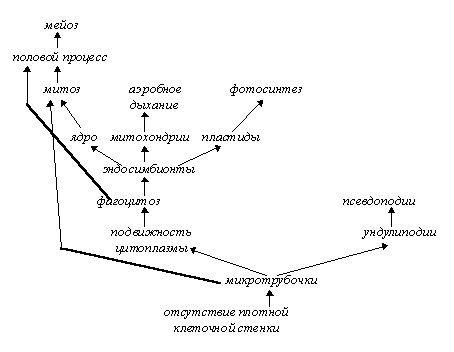
Рис. 10. Становление эукариотического типа организации.
В ходе дальнейшей эволюции у эукариот наблюдалась тенденция к диплоидизации первичных полиплоидных наборов хромосом (либо эндогенно, либо путем клеточного каннибализма), в результате чего и благодаря преадаптации в виде микротубулярных систем смог сформироваться половой процесс, а затем мейоз как способ стабилизации числа хромосом и возврата их к гаплоидному (первично-полиплоидному), причем мейоз возникал широко полифилетично во многих группах, находящихся на разных уровнях организации и в разных филумах, в отличие от митоза, который возник если и не единожды, то, во всяком случае, небольшое число раз, у некоторых наиболее примитивных групп эукариот. На этой стадии развития из известных в настоящее время организмов остались микроспоридии и так называемые архезои - группа жгутиковых и амебоидных "простейших", первично лишенных митохондрий (Diplomonada, Retortomonada, Oxymonada, некоторые другие “животные жгутиконосцы”, амебы Pelomyxa, Endamoeba) [Мирабдуллаев; Самовар, 91]. Возможно, эти организмы, как и динофлагелляты, обладают кольцевыми безгистоновыми хромосомами, так как биохимические данные свидетельствуют о том, что они отделились от остальных эукариот раньше, чем динофлагелляты, имеющие, к тому же, эндосимбионтов в виде митохондрий и пластид. Кроме того, микроспоридии характеризуются бактериальным типом рибосом, бактериальными размерами, уникальной формой митоза, плотной клеточной оболочкой, отсутствием полового размножения и эндосимбионтов и рядом других признаков [Мирабдуллаев, 89], на основании которых можно сделать вывод о том, что, возможно, микроспоридии приобрели эукариотическую организацию и митоз независимо от прочих эукариот, причем происходят они, видимо, от эубактерий, а не от организмов, родственных археям, как, по-видимому, остальные эукариоты. В этом случае ядро микроспоридий, как и ядро настоящих эукариот, приобретено либо путем мембранизации, либо в результате симбиоза с еще более мелким организмом, однако второй путь кажется нам маловероятным. Так или иначе, микроспоридий следует выделять в отдельное царство, а в случае получения подтверждений гипотезы об их независимом выходе на эукариотический уровень их следует убрать из группы, включающей архей и эукариот, и поместить в группу, к которой относятся бактерии.
Помимо микроспоридий, архезой и динофлагеллят, группой, отделившейся до разделения первичных эукариот на два наиболее крупных филума - Tubulicristata и Lamellicristata - следует считать инфузорий, так как они обладают рядом уникальных признаков (например, половым процессом в виде сложной конъюгации, не имеющей ничего общего с так называемой конъюгацией зеленых водорослей - конъюгат, или зигнематофитов, - которая, в свою очередь, схожа, скорее, с зигогамией и соматогамией настоящих грибов), а строение их крист чисто трубчатое, в отличие от крист остальных Tubulicristata, либо выглядящих как пузырек на тонкой ножке (Heterocontae и др.), либо ветвящихся и анастомизирующих [Мирабдуллаев].
Другой достаточно древней группой эукариот является эволюционная ветвь
Discocristata, включающая такие группы, как Euglenophyta, Protomonadida (Kinetoplastida), Pseudociliata (Stephanopogon sp.), солнечники Nucleariida, а также Heterolobosea (Acrasiomycota и Schizopirenida), характеризующиеся дисковидными кристами митохондрий, отсутствием полового размножения, открытым ортомитозом с сохранением ядрышка, передним положением цитостома и общностью биохимических особенностей [Мирабдуллаев]. По-видимому, Discocristata, как инфузории и динофлагелляты, независимо от Tubulicristata и Lamellicristata происходят от какой-то группы первичных эукариот, наделенных митохондриями, однако не исключена и общность их происхождения с пластинчатокристовыми, о чем свидетельствуют переднее расположение цитостома и наличие ортомитоза [Мирабдуллаев]. Впрочем, не исключено и независимое приобретение митохондрий различными группами эукариот, однако пока, видимо, нет оснований полагать, что такое явление имело место.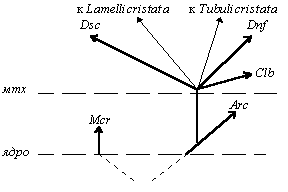
Рис. 11. Начальная эволюция эукариот: мтх - митохондрии; Mcr - микроспоридии, Arc - архезои, Clb - инфузории, Dnf - динофлагелляты, Dsc - дискокристовые.
Итак, уже до выделения двух магистральных стволов эволюции эукариот сформировались, по крайней мере, два царства организмов прокариотического типа (добавляя, в целях унификации таксономии, к названиям царств окончания
-bionta, мы назовем эти царства Bacteriobionta и Archaebionta) и 4-5 царств (в зависимости от того, выделять ли эволюционно связанных с тубуликристатами, но более примитивных динофлагеллят в отдельное царство) эукариот (Microsporidiobionta, Archaezoobionta, Ciliobionta, Discocristobionta и, возможно, Dinobionta) (см. рис. 11). Tubulicristata.В этом разделе будет кратко рассмотрена эволюция ствола
Tubulicristata, включающего несколько ветвей, в одну из которых входят гетероконтные организмы с трехразделенными трубчатыми мастигонемами и производные от них группы, а в другую - в основном, амебоидные формы либо жгутиконосцы с ветвящимися кристами и гетероконтным, но безмастигонемным жгутикованием [Мирабдуллаев]. Кроме того, обособленно здесь стоят такие группы, как Vaucheriales, совмещающие некоторые черты обеих ветвей, Stephanocontae (Oedogoniales, Siphonales) с уникальным стефаноконтным жгутикованием [Айкхорн и др., 90; Водоросли, 89; Гарибова и др., 75; Жизнь растений; Курс низших растений, 81; Лемеза, Шуханов, 94] и различные группы споровиков. Кроме строения митохондриальных крист, тубуликристаты характеризуются также преимущественно задним расположением цитостома, что отличает их как от дискокристат, так и от ламелликристат.Судя по сходству митохондрий тубуликристат с митохондриями инфузорий и динофлагеллят [Мирабдуллаев, 89] можно предположить, что трубчатое строение крист является близким к базовому, послужившему исходным материалом для других типов крист. По-видимому, уже на ранних этапах эволюции от первичных
Tubulicristata отделились высокоспециализированные Cnidosporidia, Gregarinida и Coccidiida, перешедшие к паразитическому образу жизни. Вообще, принадлежность этих групп к трубчатокристовым, как и их родственные связи, можно поставить под сомнение, так как они имеют совершенно разные жгутиковые аппараты [Догель, 80], являющиеся в большинстве крупных таксонов важным систематическим признаком. Более того, такие жгутиковые аппараты больше не встречаются среди тубуликристат, зато характерны для ламелликристат: одиночный задний жгутик грегаринид напоминает таковой у хитридиомицетов и большинства многоклеточных животных [Малахов, Реунов], а изоконтное жгутикование, имеющее место у кокцидий, характерно для зеленых водорослей и высших растений [Адрианов, 96; Айкхорн и др., 90]. В связи с этим данные группы, ранее объединяемые под названием споровиков, заднежгутиковых (каудоконтных) Spyromonas, а также книдоспоридий, мы пока что условно отнесем к Tubulicristata [Мирабдуллаев], считая все три ветви рано и независимо друг от друга отделившимися от этого ствола и происходящими от разных примитивных групп трубчатокристовых.На основании сходства жгутикового аппарата с таковым у динофлагеллят, можно предположить, что базовой группой тубуликристат является группа, характеризующаяся трубчато-пузырчатыми кристами, общностью биохимических особенностей и гетероконтным жгутикованием, причем один из жгутиков обычно имеет мастигонемы, однако некоторые из представителей этой группы имеют другой характер жгутикования, производный от этого. Исходя из данных сравнения нуклеотидных последовательностей в рибосомных РНК, к этой группе отнесены гелиофлагелляты
Ciliophryidae, солнечники Actinophryidae и группа Heterocontae, или Pedinellida, включающие бурые, золотистые, желто-зеленые, диатомовые (возможно) водоросли, празинофитов, рафидофитов, а также Oomycota, Hyphochytridiomycota, Traustochytridiomycota и Labyrinthulomycota [Мирабдуллаев], причем очевидно, что все гетероконты представляют собой единую эволюционную и систематическую группу, тогда как остальные трубчатокристовые этой ветви могли возникнуть от общих предков независимо от гетероконт. Что интересно, приобретение гетероконтами пластидных симбионтов происходило неоднократно, причем разные группы гетероконтных водорослей приобретали разных бактериальных партнеров из группы Proplastidae, обладающих различными наборами фотосинтезирующих пигментов (помимо содержащих фикобилины и хлорофилл а цианобактерий, это могли быть бактерии наподобие содержащего хлорофиллы а и b Prochloron или другие, не известные в настоящее время в свободном состоянии [Маргелис, 83], давшие начало пластидам оранжево-желтых тонов). Эти же бактерии вступали в симбиозы и с эукариотами других филумов.Вторая большая группа трубчатокристовых включает амеб
Lobosa, Chlororarchnion, акантамеб, фораминифер, настоящих солнечников Euheliozoa, Centrohelidia, настоящих слизевиков (Protostellida, Discostellida, Myxogastrea), жгутиконосцев Traumatomonadida, Pseudodendromonadida, Rhipidodendromonadida, Gyromitis, Pseudosporea, а также слопалинид (опалины и псевдоспореи), протеромонад, радиолярий [Мирабдуллаев] и, возможно, гемоспоридий и гаплоспоридий. Несмотря на явную биохимическую и цитологическую обособленность этой группы живых организмов от гетероконт, общность их происхождения несомненна, что подтверждают не только данные цитологии и биохимии, но и существование таких организмов, как Vaucheria, на разных стадиях своего жизненного цикла имеющие как мастигонемные, так и безмастигонемные жгутиковые формы [Жизнь растений; Лемеза, Шуханов, 94, стр. 88-89]. По-видимому, ваушериевые водоросли представляют собой потомков форм, переходных между первой и второй группами трубчатокристовых, или, скорее, группу, происходящую от общего предка обеих ветвей.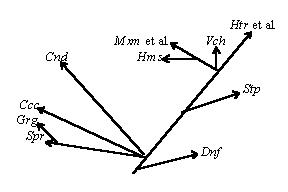
Рис. 12. Эволюция тубуликристат: Dnf - динофлагелляты, Stp - стефаноконты, Htr et al.- гетероконты и другие, Vch - ваушериевые, Mxm et al. - миксомицеты и другие, Hms - гемоспоридии, гаплоспоридии, Spr - спиромонады, Grg - грегарины, Ccc - кокцидии, Cnd - книдоспоридии.
Кроме перечисленных групп, к трубчатокристовым (в частности, к настоящим слизевикам) относят иногда плазмодиофоровых, однако имеются данные, что кристы их имеют скорее пластинчатое, чем трубчатое строение [Мирабдуллаев, 89] (см. рис. 12)
. Lamellicristata.Группа
Lamellicristata происходит либо от общего предка независимо от Tubulicristata, либо от какой-то примитивной группы Tubulicristata. Некоторые цитологические особенности, как было сказано, дают возможность также полагать, что пластинчатокристовые находятся в родстве с дискокристовыми, однако эти особенности могли возникнуть и параллельно. Помимо пластинчатого строения митохондриальных крист, эта группа характеризуется тенденцией к многоклеточности и переходу к наземному образу жизни, что, вероятно, связано с большей энергоемкостью пластинчатокристовых митохондрий по сравнению с другими формами. К этой группе относятся несколько довольно примитивных групп (хоанофлагелляты, хитридиомицеты, глаукофиты, хлорофиты, криптофиты и, возможно, гаптофиты), а также такие высокоразвитые группы, как красные водоросли, высшие растения, настоящие грибы и многоклеточные животные [Мирабдуллаев]. В этом разделе будет кратко рассмотрена эволюция пластинчатокристовых живых организмов, кроме животных, эволюция которых будет рассмотрена далее.На ранних этапах эволюции ламелликристаты разделились на две основные ветви (см. рис. 13), первая из которых имела преимущественно изоконтный тип жгутикования, причем оба жгутика располагались на переднем конце клетки, а вторая - один задний жгутик (причем двужгутиковая форма, вероятнее всего, является первичной, так как проявляется на сперматозоидной стадии у некоторых примитивных животных - представителей моноконтной ветви пластинчатокристовых [Малахов, Реунов], но стадий с задним жгутиком у изоконт и их потомков не обнаружено; другую версию, которая объясняла бы это явление, см. ниже). Кроме того, обособленно стоят среди ламелликристат криптофиты, имеющие характерный для них тип жгутикования (возможно, сестринской для них группой являются плазмодиофоровые, если верна информация о том, что кристы у них гребневидные), и группа, включающая глаукофитов и родофитов, хотя не исключено
, что последние состоят в родстве с грибами.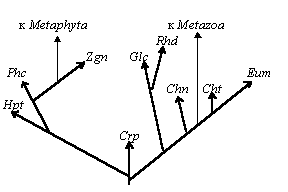
Рис. 13. Эволюция ламелликристат: Crp - криптофиты, Hpt - гаптофиты, Phc - настоящие (зеленые) водоросли, Zgn - зигнематофиты, Glc - глаукофиты, Rhd - красные водоросли, Cht - хитридиомицеты, Eum - высшие грибы, Chn - хоанофлагелляты.
Что касается первой ветви, то она представлена изоконтными зелеными водорослями и высшими растениями, а также, возможно, гаптофитами, которые хотя и приобрели пластидных симбионтов независимо от хлорофитов, но схожи с ними изоконтным жгутикованием [Жизнь растений]. Что касается зеленых водорослей, то по современным данным их делят на классы
Micromonadophyceae, Pleurastophyceae, Ulvophyceae и Chlorophyceae, которые различаются особенностями клеточного строения и метаболизма[Айкхорн и др., 90], а не уровнем организации, как это было в старой систематике.От какой-то ветви зеленых водорослей (вероятно, одноклеточных) отделилась ветвь, характеризующаяся такими признаками, как деление с образованием фрагмопласта и клеточной пластинки, с сохранением веретена деления и разрушением ядерной оболочки в начале митоза, оогамное оплодотворение, асимметрия жгутиковой стадии, поперечное деление зиготы и многие общие особенности биохимии [Адрианов, 96; Айкхорн и др., 90; Бардунов, 84; Жуковский, 64; Седова]. Эта группа дала начало двум ветвям, одна из которых в результате утраты жгутиковой стадии дала начало конъюгатам, или зигнематофитам, а другая путем повышения уровня эмбрионализации и тканевой организации, привела к возникновению высших растений. Причем, по-видимому, достижение представителями этой группы уровня собственно “высших растений” и переход к наземному образу жизни происходил неоднократно, по принципу полифилии. Фрагмопласт и клеточная пластинка являются, по-видимому, наиболее характерным признаком настоящих растений (свойственным всем представителям этой и только этой группы), на основании чего к высшим растениям, помимо высших сухопутных мохообразных и сосудистых, следует относить также харовые, колеохетовые и некоторые другие водоросли (в т.ч.
Klebsormidium), а, возможно, и зигнематофитов, для которых также характерно наличие фрагмопласта и клеточной пластинки (наличие среди зигнематофитов одноклеточных форм свидетельствует о том, что фрагмопластовые организмы - самостоятельная ветвь в эволюции растений, а не просто уровень организации).Можно умозрительно описать общего предка наземных фрагмопластовых растений. Это были многоклеточные гаплоидные водоросли с оогамным половым процессом, многоклеточными гаметангиями и зиготической редукцией, зигота которых прорастала под покровами тканей материнского организма (либо на самом теле материнского гаметофита в оокарпе, как у колеохет, либо в особых отбрасываемых почках, как у харовых). Их зооспоры и антерозоиды имели асимметричное строение и обладали двумя равными жгутиками [Айкхорн и др., 90]. Хлоропласты первично были мелкими, зернистыми, лишенными пиреноидов и возникли в результате симбиоза с бактерией типа Prochloron [Маргелис, 83], однако у некоторых специализированных групп (например,
Coleochaete, Anthoceratophyta) позднее преобразовались в крупный [Бардунов, 84]. Обитал такой организм, скорее всего, в пресных водах.От подобного организма произошли две ветви, одна из которых, более схожая с современными колеохетами и обладающая видоизмененными пластидами, перешла к гаметической редукции (при этом развивающийся на гаметофите спорофит, или диплонт, был как бы растянувшейся стадией зиготы [Петров, 74], в связи с чем стоит сформулировать положение о том, что гаплонт всегда гомологичен гаплонту, тогда как диплонт гомологичен другому диплонту только в группах с монофилетически возникшим половым процессом, как в случае с фрагмопластовыми растениями), утратила зооспоры и дала начало антоцеротовым, а другая, сохранившая примитивный тип пластид, напоминающих бактериальные клетки, и зиготическую редукцию, - всем остальным группам. При этом оставшиеся на этом уровне организмы образовали группу харовых, а те, которые, параллельно антоцеротовым, приобрели чередование многоклеточных диплонта и гаплонта, продолжили прогрессивную эволюцию и завоевали сушу.
Дальнейшая эволюция высших растений представляет собой цепочку преобразований, направленных на преобладание в жизненном цикле диплонта: диплонт не способен к самостоятельному существованию - диплонт способен к самостоятельному существованию, но преобладает гаплонт - диплонт и гаплонт схожи между собой - диплонт преобладает, но гаплонт похож на него - гаплонт выходит из споровой оболочки, но не похож на диплонт -
гаплонт не выходит из споровой оболочки, - и на разнополовость гаметофита: изоспория - физиологическая гетероспория - морфологическая гетероспория при наличие одного типа спорангиев - образование мужских и женских спорангиев - двудомность. При этом достижение одних и тех же стадий могло происходить независимо в разных группах [Адрианов, 96].Одну группу наземных высших растений образуют те, которые сохранили изоконтные двужгутиковые антерозоиды: Bryophyta, Hepatophyta, Lycopodiopsida, Selaginellopsida [Адрианов, 96; Жуковский, 64; Шляков, 75]. Монофилетически (в результате единственной мутации) от высших изоконт происходит ветвь, в которой антерозоиды приобрели многожгутиковость: Isoёtopsida, Tmesophyta, различные группы папоротников, которые могут являться как моно-, так и полифилетичной группой,
Cycadophyta, Ginkgophyta, а также группы, утратившие (скорее всего, полифилетично) жгутиковую стадию: хвойные, гнетовые и покрытосеменные [Адрианов, 96]. Что касается происхождения покрытосеменных растений, то ряд признаков (в первую очередь наличие сосудов, редукция архегониев и половой процесс, близкий к двойному оплодотворению) сближает их с гнетовыми, однако, по-видимому, и те и другие происходят от общего предка, обладавшего более примитивным строением, происходящие от которого ветви в силу общности эволюционных тенденций приобретали схожие признаки [Айкхорн и др., 90; Антонов, Боброва; Жуковский, 64]. При этом, по-видимому, и уровень собственно покрытосеменных возникал неоднократно, так что нынешние цветковые растения, скорее всего, представляют собой полифилетичную группу (см. рис. 14).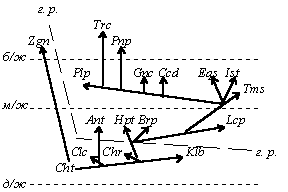
Рис. 14. Эволюция высших растений: д/ж - двужгутиковые, м/ж - многожгутиковые, б/ж - безжгутиковые; г. р. - гаметическая редукция; Cht - хетофоровые, Klb - клебсормидиум, Clc - колеохетовые, Ant - антоцеротовые, Chr - харовые, Hpt - печеночники, Brp - мохообразные, Lcp - плауновидные, Tms - тмезоптеровые, Ist - изоэтовые, Eqs - хвощи, Plp - папоротникообразные, Ccd - цикадовые, Gn
c - гинкговые, Pnp - хвойные, Trc - трахеофиты (гнетовые и покрытосеменные).Другой ствол
Lamellicristata, жгутиковые стадии представителей которого в большинстве своем каудоконтны, представлен в основном ветвями, завершающимися настоящими грибами и высшими животными. От этого же ствола, причем, скорее всего, от ветви, приведшей к возникновению грибов (такой вывод можно сделать исходя из принципа эволюционных тенденций), происходят, вероятно, глаукофиты и красные водоросли [Мирабдуллаев].Что касается предковых форм этого ствола, то они, вероятно, имели в своем жизненном цикле не только одножгутиковые, но и двужгутиковые стадии. Наиболее рано специализировавшаяся группа этой ветви утратила двужгутиковую стадию и дала начало хитридиомицетам, от которых, в свою очередь, произошли зигомицеты, аскомицеты и базидиомицеты. Интересная деталь в эволюции грибов - приобретение многими из них жизненной формы дрожжей, причем такая жизненная форма встречается у разных типов настоящих грибов. Дрожжи интересны тем, что способны вторично утрачивать митохондрии и переходить к анаэробиозу, однако генетический материал митохондрий сохраняется, и митохондрии могут формироваться вновь [Мирабдуллаев]. Что же касается дейтеромицетов или, как их называют, “несовершенных грибов”, то в действительности они представляют собой утративших половой процесс аскомицетов или базидиомицетов и таким образом, несомненно, являются искусственной группой.
Сохранившая в своем жизненном цикле примитивную двужгутиковую стадию ветвь, как следует из принципа тупиковости специализации, смогла продолжить свою эволюцию более широко и привела к возникновению царства животных.
3. 3. 6. Горизонтальная эволюция беспозвоночных.
Наиболее примитивными из ныне известных животными (с эволюционной, а не уровневой или экологической точки зрения) можно назвать так называемых воротничковых жгутиконосцев (Choanoflagellata) [Беклемишев, 79, стр. 50; Иванов, 68], однако считать современных хоанофлагеллят предками многоклеточных, безусловно, нельзя. По-видимому, настоящие предки многоклеточных еще не утратили двужгутиковую стадию, так как некоторые из низших многоклеточных (турбеллярии, гастротрихи и, возможно, связанные с ними группы, относимые к плоским и круглым червям) обладают двужгутиковыми сперматозоидами [Малахов, Реунов]. Впрочем, мы не исключаем и возможной вторичности двужгутиковой формы у животных. Действительно, явное филогенетическое родство групп со сперматозоидами, обладающими двумя жгутиками, может говорить о том, что второй жгутик появился в результате единственной мутации у предков турбеллярий, гастротрих и родственных им групп. Эти предки образовывали шаровидные колонии, состоящие из небольшого числа клеток и плавающие за счет выставленных наружу жгутиков, и имели слабую дифференцировку на передний и задний полюс. Бластопорального отверстия и бластоцеля они не имели, так как колонии были небольшими, и все клетки контактировали с окружающей средой и, внутренними концами, друг с другом, поэтому все клетки питались самостоятельно. Так как этот первопредок животных рекапитулируется морулой или стадией нескольких бластомеров, то его можно назвать мореллой. Судя по многообразию способов определения пола у животных и, следовательно, по причине полифилетичного возникновения у них полового процесса, можно сделать вывод, что способностью к половому размножению общий морелла (а также его прямые потомки - фагоцителлы всех уровней) не обладала. Вероятно, она обладала способностью к агаметическому (первично-партеногенетическому) размножению, агамета делилась палинтомически, а первая борозда дробления была врезающейся, а не кольцевой, как у большинства высших животных [Вагин, 68].
Далее эволюция животных шла в горизонтальном и вертикальном направлениях (см. п. 3. 2. 3., рис. 5). Что касается горизонтального, то оно является фактически эволюцией свободноплавающих водных неотенических личинок, которых называют также фагоцителлами. От различных фагоцителл в течение достаточно короткого времени, но в разных направлениях, отделялись вертикальные ветви, которые, в свою очередь, могли проходить стадии, соответствующие фагоцителлам более высокого порядка, чем их предковые [Беклемишев, 79, стр. 183]. Это затрудняет выявление происхождения разных групп животных от той или иной фагоцителлы: обычно можно сказать только, что предковая фагоцителла была уровнем не выше такой-то (соответствующей по уровню организации изучаемой группе), - поэтому сперва отдельно будет рассмотрена горизонтальная эволюция беспозвоночных, или эволюция фагоцителл.
Рис. 15. Эволюция фагоцителл (дублирование уровней организации в ходе разных направлений эволюции не учтены).
Фагоцителловые стадии обычно рекапитулируются в эмбриональном развитии животных, причем, по-видимому, фагоцителл было гораздо больше, чем принято считать (см. рис. 15). Наиболее примитивной фагоцителлой, которую можно назвать бластеллой, так как ее рекапитулирует бластула, следует считать шаровидную колонию примитивных хоанофлагеллят, не имеющую еще, по-видимому, ротового отверстия. Бластелла имела единственный эпителий, во внутреннюю же среду эпителиальные клетки выходили только для питания и агаметического размножения, приобретая для этого амебоидную форму и временно утрачивая жгутик.
В ходе дальнейшей эволюции бластоцель у фагоцителлы заполнился неэпителизированной клеточной массой, служащей для поглощения попадающих в первичную пищеварительную полость питательных частиц – профагоцитобластом, - в связи с чем возникла необходимость в формировании бластопорального отверстия. Вероятно, клетки профагоцитобласта первоначально не постоянно занимали свое место внутри организма, а время от времени мультиполярно переходили в бластоцель из наружного эпителия, однако, поскольку слой этот, в целом, присутствовал постоянно, эту фагоцителлу можно назвать паренхимеллой.
Позднее фагоцитобласт стал образовываться в результате сперва униполярной иммиграции, а затем – инвагинации (как это следует из принципа экономичности развития), что привело к возникновению гастреллы – фагоцителлы с двумя эпителиями (в другой ветви, происходящей, видимо, от бластеллы или, скорее, от мореллы, фагоцитобласт стал образовываться путем деляминации, что привело к возникновению гастреллярной фагоцителлы быстрее, чем через стадию с униполярной иммиграцией; впрочем, гаструляция могла также идти и другими путями). Далее между двумя эпителиями мультииммиграционно либо путем вселения одной или нескольких клеток возник третий слой, который изначально не был эпителизованным [Беклемишев, 79, стр. 38; Зусман, 73]. Стадию с двумя эпителиями – фагоцитобластом и кинетобластом – и неэпителизованной клеточной массой между ними можно назвать постгастреллой.
У постгастреллы, возникшей путем мультиполярной иммиграции (либо даже у гастреллы) в первичной кишке могли происходить впячивания и образовываться своеобразные карманы (первоначально, вероятно, для увеличения площади всасывания). Постепенно все более отшнуровываясь от бластоцеля, они привели к образованию целомов у так называемых вторичноротых и родственных им групп, происходящих от диплевреллы – фагоцителлы с инвагинационно (энтероцельно) возникающим третьим эпителием [Беклемишев, 79, стр. 173].
Целомы могли возникать и другими путями. Так, у ветви, приведшей к возникновению трохофорных животных (фагоцителла-трохофорелла), они образовывались из вселившихся в межэпителиальную прослойку (“мезоглею”) отдельных клеток. Кроме того, не исключено, что у некоторых организмов сперва происходило рассасывание у постгастреллы неэпителизированной внутренней массы, а образовавшиеся полости позднее приобретали эпителий (сходный, в какой-то степени, процесс мы можем наблюдать при формировании кровеносных сосудов у немертин).
Таким образом, даже в эволюции наиболее примитивных животных - фагоцителл - можно обнаружить определенную ветвистость. При этом, как уже было сказано, фагоцителлы, эволюционирующие в том или ином направлении, могли неоднократно и независимо друг от друга проходить одни и те же стадии. Что же касается других животных, появлявшихся в результате вертикальной эволюции от различных фагоцителл, то они возникали обычно из фагоцителл, перешедших к бентосной форме существования. При этом особенности организации и строения этих животных существенно различались в зависимости от того, на какой полюс оседала
фагоцителлообразная личинка [Беклемишев, 79, стр. 183], а также от способа возникновения полового процесса и уровня организации, на котором он приобретался.3. 3. 7. Вертикальная эволюция беспозвоночных.
Наиболее рано к вертикальной эволюции перешли ветви, приведшие к возникновению некоторых низших червей и пластинчатых, которые либо остановились на стадии паренхимеллы, либо продвинулись далее по пути эпителизации, но явно эволюционно зависимы от паренхимеллообразных форм. Характерной особенностью червей этой ветви - турбеллярий, гастротрихов и, возможно, некоторых других родственных им плоских и круглых червей (причем плоские и круглые черви – группы явно полифилетичные, подлежащие разделению по разным эволюционным ветвям и, следовательно, по разным таксонам) – является наличие у них двужгутиковых сперматозоидов [Малахов, Реунов], рекапитулирующих, возможно, двужгутикового пластинчатокристового предка (о другой трактовке этого феномена см. выше). Черви этой группы происходили от фагоцителлы, осевшей на брюшную сторону [Беклемишев, 79, стр. 184], в результате чего рот у примитивных турбеллярий находится на заднем конце, а плакозои осели, видимо, еще раньше, до образования ротового отверстия, открывающего путь к фагоцитобласту.
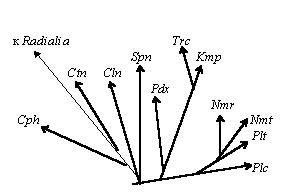
Рис. 16. Эволюция животных: Plc - пластинчатые , Plt - плоские черви, Nmt - настоящие круглые черви, Nmr - немертины, Kmp - камптозои, Trc - высшие трохофорные, Pdx - актинотрохные, Spn - губки, Cln - кишечнополостные, Cph - головохоботные, Ctn - гребневики.
На этом же уровне отделилась другая, более крупная ветвь, полностью утратившая двужгутиковую стадию (впрочем, пример грибов показывает, что двужгутиковая стадия у ламелликристат могла утрачиваться неоднократно в разных эволюционных ветвях). Наиболее примитивными из этих животных, происходящими от фагоцителл уровня от паренхимеллы до гастреллы, являются губки (не исключено, что губки по своему происхождению по крайней мере дифилетичны, так как развитие у них может идти либо через амфибластулу, либо через паренхимулу [Беклемишев, 79, стр. 39; Догель, 80; Захваткин, 49; Иванов, 68], однако их противопоставление остальным многоклеточным животным на основании экскурвации зародышевых листков вряд ли обосновано), кишечнополостные и, вероятно, некоторые доцеломические черви с одножгутиковыми сперматозоидами. При этом губки и кишечнополостные осели на аборальный полюс, а черви, как и в предыдущей группе, на спинную либо брюшную сторону
[Беклемишев, 79, стр. 184].От трохофореллы сперва отделились осевшие на аборальный полюс камптозои, не сформировавшие еще целомов, и целомические сипункулиды (а также, возможно, какие-то родственные им черви), а позднее – группы, монофилетично или полифилетично сформировавшие целомы и осевшие на оральный полюс (в связи с этим и рот, и анус у них возникают на месте частично затянувшегося бластопора). При этом в зависимости от того, сколько целомических мономеров изначально образовывалось, широкой полифилией возникали различные группы олигомерных и полимерных трохофорных организмов [Беклемишев, 79, стр. 129, 139].
Олигомерные трохофорные животные целиком состоят из ларвального тела, не образуя постларвальных сегментов [Беклемишев, 79, стр. 129]. По этому признаку можно сказать с известной долей приблизительности, что взрослые олигомерные животные гомологичны головному отделу животных полимерных. Наиболее примитивными из ныне известных олигомерных трохофорных являются динофилиды и схожие с ними, но уже приобретшие признаки типичных моллюсков, моноплакофоры [Беклемишев, 79, стр. 116]. Кроме того, олигомерными являются некоторые другие червеобразные животные – эхиуриды, сипункулиды и мизостомиды, - некоторые группы, ранее причисляемые к членистоногим (тихоходки, пятиустки), а также моллюски, причем все они, видимо, происходили от олигомерной трохофореллы независимо друг от друга и от полимерных форм [Беклемишев, 79, стр. 139]. Что касается моллюсков, то обычно их делят на две большие группы: либо на Peltophora и Cochleophora, либо на Amphineura и Conchiphora, - однако тот факт, что пластинчатожаберные моллюски сочетают признаки Cochleophora и Amphineura, позволяет предположить, что имело место более широкое ветвление ствола моллюсков [Беклемишев, 79, стр. 115].
Полимерные трохофорные, образующие во взрослой стадии постларвальные сегменты, представлены олигохетами, полихетами, пиявками, онихофорами и различными группами членистоногих. При этом различные кольчатые черви и даже различные полихеты (а также, возможно, погонофоры, которые, однако, могут принадлежать и к ветви Radialia) происходят, видимо, от разных трохофорелл, так как число ларвальных сегментов у них может различаться [Вагин, 68]. Онихофор ряд признаков сближает с тихоходками и пятиустками, так что, очевидно, они происходят от общих с ними олигомерных предков, самостоятельно приобретя метамерию [Беклемишев, 79, стр. 142]. Прежнее объединение трахейнодышащих и ракообразных в одну группу и противопоставление их хелицеровых, скорее всего, неверно: так как ракообразные образуют 2 ларвальных сегмента, а трахейнодышащие, хелицеровые, пантоподы и трилобиты – 4 [Беклемишев, 79, стр. 147, 152, 160], то либо противопоставление среди членистоногих противоположно традиционному, либо четырехсегментное строение сформировано у разных групп по-разному, но, в любом случае, трахейнодышащие, скорее, родственны хелицеровым и пантоподам, нежели ракообразным, причем типы организации, свойственные представителям этой группы, могли возникнуть параллельно у нескольких родственных предковых форм, о чем косвенно свидетельствует уже упомянутая множественность типов определения пола у насекомых. Все это говорит о том, что разделение трохофорных животных на олигомерных и полимерных носит уровневый, градовый характер, так как олигомерные животные – группа парафилетическая, а полимерные – полифилетическая, и поэтому необходимо уточнение их классификации.
Трохофорные животные и некоторые низшие черви, круг которых очертить пока довольно трудно за недостатком данных [Богомолов, 68], образуют эволюционный ствол Spiralia, так как они имеют преимущественно спиральный тип дробления или типы, вытекающие из спирального [Адрианов, Малахов, 95].
От примитивной трохофореллы или от более примитивной фагоцителлы происходят, вероятно, и актинотрохные животные (форониды и мшанки), осевшие, как и трохофорные, на оральный полюс. Поскольку характер дробления у них не устоявшийся (может быть и радиальным, и спиральным), их происхождение сомнительно, но, вероятно, они являются стволом, не зависящим ни от Spiralia, ни от Radialia [Адрианов, Малахов, 95; Беклемишев, 79, стр. 168].
Radialia.Остальных многоклеточных с большей или меньшей, но довольно высокой, степенью вероятности можно отнести к
Radialia, так как дробление у них радиально или сводимо к нему. Radialia происходят от диплевреллы, имеющей радиальный тип дробления и напоминающей гребневика. По некоторым данным, наиболее примитивными из Radialia являются черви, объединяемые иногда в группу головохоботных: приапулиды, киноринхи, волосатики, лорициферы, а также, возможно, скребни и коловратки. Действительно, некоторые из них имеют дробление, близкое к радиальному, и ряд других, общих с Radialia, признаков [Адрианов, Малахов, 95]. Зачастую к примитивным радиальным относят и погонофор [Адрианов, Малахов, 95; Беклемишев, 79, стр. 173; Догель, 80], но в последнее время их пытаются вновь вернуть в группу полихет.Собственно радиальными являются такие группы организмов, как плеченогие, щетинкочелюстные, иглокожие, полухордовые и различные группы хордовых [Адрианов, Малахов, 95]. Наиболее рано среди них выделились плеченогие, так как они имеют еще неустоявщийся тип образования целомических мешков, тогда как у всех остальных целомических радиальных (щетинкочелюстных и так называемых вторичноротых) они закладываются путем отшнуровывания от кишки [Беклемишев, 79, стр. 171]. Поскольку у хетогнат изначально закладывается две пары целомов, а у вторичноротых - три (иногда они вторично делятся на метамеры), можно предположить, что возникновение целомов у этих двух групп происходило независимо [Беклемишев, 79, стр. 171, 173]. Оседание же и у плеченогих, и у щетинкочелюстных (вернее, у их донных предков, так как в планктон они перешли, скорее всего, вторично) происходило, по-видимому, на брюшную поверхность [Беклемишев, 79, стр. 184].
К собственно вторичноротым, имеющим изначально три пары целомов, относятся иглокожие, полухордовые и хордовые (см. рис. 17). Однако, по-видимому, придание вторичноротым таксономического ранга не обосновано, так как явление вторичноротости в той или иной степени может достигаться и представителями других эволюционных ветвей [Адрианов, Малахов, 95].
Эволюция иглокожих весьма сложна, однако, поскольку эта группа сильно специализировалась в своем развитии, о них следует только сказать, что они происходят от диплевреллы, осевшей на аборальный (по другой версии - на оральный) полюс [Беклемишев, 79]. В качестве группы, родственной предку иглокожих, полухордовых и хордовых, можно назвать гребневиков. Гребневики, по-видимому, происходят от организмов, напоминающих диплеврулу “вторичноротых” и, следовательно, диплевреллу. Они имеют схожий с радиальным (бирадиальный) тип дробления, а их гастроваскулярный аппарат возникает энтероцельно, как и целомы высших радиальных (иглокожих, полухордовых, хордовых). В любом случае, гребневики – группа животных, уникальная в том плане, что является первично-планктонной и, таким образом, фактически осталась на уровне первичной неотенической фагоцителлы типа диплевреллы. Те из гребневиков, которые ведут в настоящее время донный образ жизни, осели на оральный полюс [Беклемишев, 79, стр. 185].
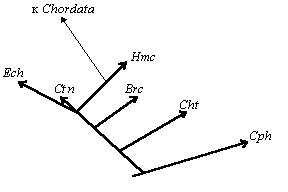
Рис. 17. Эволюция радиальных: Cph - головохоботные, Cht - щетинкочелюстные, Brc - плеченогие, Ctn - гребневики, Ech - иглокожие, Hmc - полухордовые.
Диплевреллы, приведшие к возникновению полухордовых и хордовых, осели на брюшную либо на боковую поверхность [Беклемишев, 79, стр. 185]. Одни из них (кишечнодышащие) перешли к донному роющему образу жизни, другие (крыложаберные и туникаты) - к прикрепленному. При этом туникаты сформировали вторичную расселительную личинку, передвигающуюся за счет мышечной активности хвостообразного отростка. Эта форма в результате неотенического развития стала исходной для высших форм хордовых - позвоночных [Жизнь животных; Наумов, Карташев, 79].
Сохранившейся до наших дней группой хордовых, близкой к личинке оболочников, являются ланцетники. От подобных им организмов происходят многочисленные группы рыбообразных животных, причем, по-видимому, полифилия возникновения различных групп первично-водных позвоночных гораздо шире, чем это обычно представляется. Прежде всего, обособленную позицию среди современных позвоночных занимают две группы бесчелюстных - непарноноздревые (миноги) и миксины, причем обе эти группы ведут образ жизни, близкий к паразитическому, а потому сильно специализировались, и не ясно, действительно ли они близко родственны друг другу, или же их тип организации приобретен ими независимо.
Очень сложна филогенетическая классификация рыб. Во всяком случае, очевидно, что прежнее деление их на хрящевых и костных носит градальный, уровневый характер, а не отражает реальные родственные связи. Особенности продуктов обмена, строения скелета (разумеется, не степени его окостенения, а, например, различия в соотношении базалий и радиалий в скелетах конечностей), отдельные эволюционные тенденции и ряд других признаков [Наумов, Карташев, 79] позволяют противопоставить группу костистых рыб (включающую также, возможно, костных ганоидов) группе, включающей поперечноротых (пластинчатожаберных и осетрообразных), химер, многоперов, двоякодышащих, кистеперых рыб и наземных позвоночных (см. рис. 18)
.Что касается наземных позвоночных, то происходят они, несомненно, от рипидистиевых кистеперых рыб [Наумов, Карташев, 79]. Однако, по всей видимости, рипидистиевые как группа имели тенденцию к выходу на сушу и потому делали это неоднократно. Скорее всего, даже современные наземные позвоночные происходят от разных родственных друг другу рипидистиевых, о чем свидетельствует разнонаправленная примитивность древних групп разных ветвей эволюции (например, амфицельные позвонки клювоголовых, древних синапсид и представителей других групп [Наумов, Карташев, 79, т. 2, стр. 31]). По-видимому, от одного или нескольких (2-3) видов кистеперых происходят современные амфибии; от древних амфибоидов происходит ветвь синапсид, приведшая к возникновению млекопитающих; непосредственно от рипидистиевых, без прохождения амфибоидной стадии, но полифилетично или монофилетично, происходят все другие рептилии. Если допустить монофилию остальных рептилий (анапсид, диапсид и родственных им групп), то первичным следует считать тип организации анапсид. От них (также моно- или полифилетично) происходят различные мезозойские водные ящеры (мезозавры, ихтиозавры, плезиозавры), составляющие отдельные подклассы рептилий, а также лепидозавры и архозавры [Наумов, Карташев, 79, т. 2, стр. 13]. Птицы представляют собой всего лишь сильно специализированную группу архозавров (различные псевдозухии независимо приобретали различные птичьи черты), не более и не менее уникальную, чем, например, птерозавры (которые, по некоторым данным, также были теплокровными и проявляли заботу о потомстве), поэтому их традиционное выделение не носит эволюционный характер (если предположить, что до наших дней дожили бы птерозавры, то их, видимо, тоже выделяли бы отдельным классом).
Как было сказано, синапсиды возникли от древних амфибоидов, которые происходят либо от тех же рипидистий, что все или некоторые современные амфибии, либо независимо от них, но с прохождением через амфибоидную стадию. В любом случае, они не имеют ничего общего с настоящими рептилиями, и название “зверозубые ящеры”, применяемое ко многим древним синапсидам, не совсем уместно. Разные группы тероморфных синапсид самостоятельно приобретали маммальные черты, поэтому не исключено, что монотрематы, некоторые древние млекопитающие, метатерии и эутерии могли достигнуть этого уровня параллельно, однако данных для того, чтобы прийти к тому или иному выводу на этот счет, недостаточно.
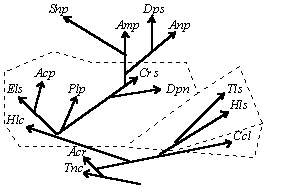
Рис. 18. Эволюция хордовых: Tnc - оболочники, Acr - бесчерепные, Ccl - круглоротые, Hls - костные ганоиды, Tls - костистые рыбы, Hlc - цельноголовые, Els - щележаберные, Acp - осетрообразные, Plp - многоперы, Dpn - двоякодышащие, Crs - кистеперые, Amp - амфибоиды, Snp - синапсиды, Anp - анапсиды, Dps - диапсиды (пунктирной линией обведены рыбообразные).
Рыбообразные и амфибоиды не имеют еще постоянного типа определения пола: пол у некоторых представителей примитивных групп позвоночных может меняться в течение жизни [Медведев, 76]. У синапсид имеет место определение пола по гомозиготности самок, причем половой механизм сформировался после становления собственно млекопитающих, о чем свидетельствует наличие у самцов рудиментарных млечных желез, а у диапсид сформировалось определение пола по гомозиготности самцов, поэтому выяснение способа определения пола у анапсид позволит ответить на
вопрос, от одной ли группы рипидистий ведут начало анапсиды и диапсиды.1. Жизнь можно определить как способ взаимодействия материи, энергии и информации, характеризующийся способностью системы к самовоспроизведению по единой информационной матрице (информационный критерий), обмену веществом, энергией и информацией с окружающей средой (экологический критерий), а также индивидуальному и историческому развитию вследствие случайных или закономерных изменений в информационной структуре (эволюционный критерий); эти три критерия являются необходимыми и достаточными для понимания сущности жизни и введение четвертого, химического критерия (белковая, или нуклеопротеиновая природа) только неоправданно сужает поле биологических исследований, не давая потенциальной возможности для изучения небелковых форм жизни.
2. Химические гипотезы происхождения жизни на Земле сталкиваются с рядом противоречий (в том числе типа вопроса о первичности курицы или яйца), тогда как получают подтверждения гипотезы космического происхождения жизни на Земле, что, однако, только отдаляет во времени и пространстве процесс ее зарождения.
3. По-видимому, некогда Земля подверглась бомбардировке своеобразными “витоспорами”, содержащими нуклеиновую кислоту (скорее всего, РНК) и взаимосвязанную с ней белковую или липопротеиновую оболочку.
4. Все “волшебные аминокислоты” происходят от единственной левоасимметричной аминокислоты, а все нуклеотиды - от единственного нуклеотида или же от их комплиментарной пары.
5. Первичные комплексы нуклеиновой кислоты и аминокислот могли напоминать современные тРНК.
6. Первоначально кодоны имели вид нуклеотидных дуплетов, а третий нуклеотид служил разделителем между кодонами. На момент возникновения современного (триплетного) генетического кода белки состояли из 7-9 типов аминокислот.
7. Клеточные формы жизни возникли благодаря симбиозу витоспор с биохимически возникшими на Земле коацерватоподобными образованьями.
8. Соединение витоспор, обладающих информационным критерием жизни, с коацерватами, обладающими экологическим критерием жизни, привело к развертыванию естественной ортогенетической “программы эволюции” и, таким образом, к появлению эволюционного критерия жизни.
9. Происхождение вирусов, очевидно, не только широко полифилетично, но и шло различными путями, среди которых наиболее вероятны возникновение вирусных частиц из витоспор, сохранившихся в результате перехода к генетическому паразитизму, автономизация участков ядерных и органоидных нуклеиновых кислот и редукция прокариот (типа риккетсий и хламидий) и прокариотических эндосимбионтов эукариот.
10. Вирусы можно разделить на несколько царств по путям их происхождения, характеру генетического материала и стратегии его реализации.
11. Вироиды и прионы являются, по-видимому, последовательными ступенями редукции витоспор.
12. Важнейшим моментом в формировании эукариотического типа ядра у ветви, близкой к археям, является приобретение (вероятно, в результате симбиоза со спироплазмоподобными бактериями) микротубулярных структур, явившееся преадаптацией для приобретения эндосимбионтов, псевдоподиального и жгутикового типа движения, митоза, мейоза, полового процесса и т.д.
13. В процессе симбиогенного формирования эукариотического типа организации могли участвовать не только археи и бактерии, но и сохранившиеся в “первичном бульоне” витоспоры и, возможно, какие-либо предбиологические системы, возникшие на Земле химическим путем.
14. Микроспоридии, по-видимому, вышли на эукариотический уровень организации независимо от других эукариот и происходят, вероятно, от эубактериальной ветви прокариот, в связи с чем всех клеточных организмов следует разделить на две ветви, одна из которых включает бактерий и микроспоридий, а другая - архей и эукариот.
15. Наиболее крупномасштабная систематика эукариот связана с типом митохондриальных крист. На основании этого признака можно выделить такие ветви, как Archaezoa, Discocristata, Tubulicristata, Lamellicristata. Кроме того, особняком стоят такие группы, как инфузории, динофлагелляты, споровики и некоторые другие.
16. К пластинчатокристовым организмам относятся наиболее высокоорганизованные живые организмы, имеющие тенденции к многоклеточности и выходу на сушу, что, по-видимому, обусловлено большей энергетической емкостью гребневидных (пластинчатых) крист по сравнению с другими типами.
17. Пластинчатокристовые разделены на два магистральных ствола, один из которых завершается цветковыми растениями, а другой - позвоночными животными.
18. Основная отличительная черта высших растений - клеточное деление с образованием фрагмопласта и клеточной пластинки. На основании этого признака к высшим растениям можно отнести, помимо наземных растений, харовые, колеохетовые и некоторые другие водоросли, а также, возможно, зигнематофитов.
19. Гаплонт всегда гомологичен гаплонту, тогда как диплонт гомологичен другому диплонту только в группах с монофилетически возникшим половым процессом.
20. Выход на уровень покрытосеменных происходил неоднократно в ветви, родственной гнетовым.
21. Эволюцию животных можно разделить на два направления - горизонтальное (эволюция фагоцителл) и вертикальное, причем фагоцителлы в своей эволюции прошли несколько стадий (морелла - бластелла - паренхимелла - гастрелла - постгастрелла - диплеврелла, трохофорелла или другая форма), - а вертикальная эволюция частично дублировала горизонтальную.
22. Базовым в систематике многоклеточных животных является разделение на Spiralia и Radialia по типу дробления зиготы.
23. В систематике рыбообразных позвоночных следует (на основании строения скелета плавников, путей возникновения плавательного пузыря, конечных продуктов азотистого обмена и некоторых других признаков) противопоставлять костистых рыб и, возможно, костных ганоидов всем остальным рыбам и наземным позвоночным.
24. Выход рипидистиевых кистеперов на сушу происходил, очевидно, неоднократно, что полифилетично привело к возникновению разных групп амфибоидов и рептилий.
25. Разные типы определения пола у разных групп позвоночных свидетельствуют о том, что бисексуальность в них возникала независимо.
1. Слабо систематизированные тезисы, касающиеся эволюционного процесса, систематизирования биологических объектов и действия умозрительного метода (принципы умозрительного метода, или ПУМ), следует учитывать при рассмотрении гипотез, имеющих отношение к эволюционной систематике и теории эволюции.
2. Процесс исторического развития жизни на Земле можно разделить по уровням организации жизни на онтогенез (индивидуальное развитие отдельных организмов), филогенез (историческое развитие систематических групп живых организмов) и сингенез (эволюция экосистем и биосферы).
3. Филогенез можно разделить на три уровня, отличающихся основными действующими на них механизмами: микроэволюцию, мезоэволюцию и мегаэволюцию. На всех уровнях имеет место множественность механизмов, однако на каждом уровне в действии этих механизмов можно проследить определенные закономерности.
4. Основные пути эволюционного развития жизни заложены в самой природе жизни подобно тому, как это имеет место на более низких уровнях организации материи.
5. Изменения, происходящие на Земле на всех уровнях, находятся в тесной зависимости от астрофизических процессов. Взаимодействие более масштабных и менее масштабных явлений происходит по принципу “эволюционного отражения”.
6. Жизнь можно определить как способ взаимодействия материи, энергии и информации, характеризующийся способностью системы к самовоспроизведению по единой информационной матрице (информационный критерий), обмену веществом, энергией и информацией с окружающей средой (экологический критерий), а также индивидуальному и историческому развитию вследствие случайных или закономерных изменений в информационной структуре (эволюционный критерий); эти три критерия являются необходимыми и достаточными для понимания сущности жизни и введение четвертого, химического критерия (белковая, или нуклеопротеиновая природа) только неоправданно сужает поле биологических исследований, не давая потенциальной возможности для изучения небелковых форм жизни.
7. Жизнь на Земле возникла в результате объединения космических витоспор, содержащих нуклеиновые кислоты и обладающих информационным критерием жизни, и предбиологических систем (в том числе коацерватоподобных), образовавшихся химическим путем на Земле и обладающих экологическим критерием жизни. В результате этого симбиоза была запущена “программа эволюции” (возник эволюционный критерий жизни).
8. Генетический код формировался в результате постепенного химического видоизменения исходных нуклеотидов и аминокислот. Триплетный генетический код возник из дуплетного.
9. Различные группы вирусов, вероятно, происходят от автономизировавших клеточных структур (в том числе симбиогенных), перешедших к генному паразитизму витоспор или регрессировавших прокариот (последняя версия, в частности, может оказаться верной для поксвирусов), причем основным критерием их классификации является характер наследственного материала и стратегия его реализации.
10. Важнейшим моментом в формировании эукариотического типа ядра у ветви, близкой к археям, является приобретение (вероятно, в результате симбиоза со спироплазмоподобными бактериями) микротубулярных структур, явившееся преадаптацией для приобретения эндосимбионтов, псевдоподиального и жгутикового типа движения, митоза, мейоза, полового процесса и т.д.
11. Можно выделить, по крайней мере, 5-8 царств вирусов, 2-4 царства прокариот и 12-17 царств эукариот на основании биохимических и структурных особенностей. Наиболее крупномасштабная систематика эукариот основывается на строении митохондриальных крист и коррелирующих с этим признаком особенностях.
12. Эволюцию животных можно разделить на два направления - горизонтальное (эволюция фагоцителл) и вертикальное, причем фагоцителлы в своей эволюции прошли несколько стадий (морелла - бластелла - паренхимелла - гастрелла - постгастрелла - диплеврелла, трохофорелла или другая форма), - а вертикальная эволюция частично дублировала горизонтальную.
1. Современная эволюционная наука переживает кризис, выходу из которого может способствовать гармоничное взаимодействие между методами точных, естественных и гуманитарных наук.
2. К настоящему времени назрела необходимость “нового синтеза” эволюционной теории, основанного на примирении дарвинистских, ортогенетических и других концепций, существующих в теории эволюции.
3. Умозрительный метод, главным критерием достоверности в котором является взаимная логическая и фактическая непротиворечивость данных, зачастую является единственным доступным методом при изучении эволюции на надвидовом уровне.
4. Некоторые закономерности эволюционного процесса, а также представления, касающиеся смежных областей знаний, можно сформулировать в виде принципов умозрительного метода (ПУМ), которые следует учитывать при умозрительном анализе эволюционной систематики и теории эволюции.
5. Историческое развитие жизни на Земле по уровням организации делится на онто-, фило- и сингенез. В свою очередь, филогенез, представляющий собой основной объект теории эволюции, можно разделить на три уровня (микро-, мезо- и мегаэволюция), различающихся между собой действующими на них эволюционными механизмами и закономерностями (не исключена также возможность более детального дробления эволюционного процесса). Уровни эти не вытекают один из другого, но взаимно влияют друг на друга, и выяснение того, продуктом какого процесса является то или иное эволюционное преобразование, поможет в дальнейшем унифицировать эволюционную систематику.
6. Эволюция в целом имеет ортогенетический характер, что связано с универсальными свойствами материи.
7. Жизнь можно определить как способ взаимодействия материи, энергии и информации, характеризующийся информационным, экологическим и эволюционным критерием. Согласно приведенной в данной работе космическо-теллурической гипотезе происхождения жизни на Земле, информационный критерий унаследован от космических витоспор, экологический - от автохтонных предбиологических систем, возникших химическим путем, а эволюционный возник в результате взаимодействия первых двух.
8. В ходе эволюции сформировалось не менее 5-8 царств вирусов, 2-4 царств прокариот и 12-17 царств эукариот, отличающихся биохимическими и структурными особенностями (деление на царства представлено в приложении №1).
В данной работе нами были рассмотрены некоторые аспекты применения умозрительного метода к таким общебиологическим дисциплинам, как эволюционная систематика и теория эволюции.
Основным вкладом, внесенным нами в эволюционную теоретику, является формулирование принципов умозрительного метода в эволюционной науке, разделение макроэволюционного процесса на мезо- и мегаэволюцию, описание и систематизация основных механизмов надвидовой эволюции, введение понятий о мегаэволюционной детонации, эволюционном зеркале в сингенетических процессах, гомофилии и гетерофилии в эволюции вирусов и трех необходимых и достаточных критериях жизни, формулировка космическо-теллурической (расширенной симбиогенной) гипотезы возникновения жизни на Земле, описание становления эукариотического типа организации в связи с возникновением микротубулярного аппарата и построение предположительной общей картины эволюционного процесса на Земле, а также некоторые другие, более частные моменты. Также составлены систематические таблицы царств (с диагностикой и представителями), типов и классов живых организмов.
Данная работа носит обзорный характер. Ближайшим перспективным направлением при рассмотрении вопросов эволюционной науки, которым мы намерены заниматься в случае продолжения научной деятельности на базе аспирантуры, является приложение умозрительного анализа к неэволюционным формам систематики (экологическая и морфологическая систематика, систематика уровней организации) и составление на этой основе периодической системы типов организации (тема работы - “Неэволюционная систематика и вопрос о периодической системе форм жизни” - служит развитием затронутой в нашей нынешней работе темы об объективной эволюционной системе живых организмов; прежде всего работа будет вестись по наиболее глобальным подразделениям живой природы, а также по систематике животных), а также инвентаризация, анализ и систематизация всех доказанных и предполагаемых механизмов эволюционного процесса (предполагаемая тема - “Общая система эволюционных механизмов” - развивает вопрос о механизмах эволюции, затронутый здесь). Кроме того, в последнее время начала разрабатываться нами тема “Этапы и механизмы формирования современного генетического кода”. Работа в этом направлении должна послужить созданию путем нового синтеза общей эволюционной теории и современной, по-настоящему филогенетической, систематики.
ПРИЛОЖЕНИЕ 1. Диагностика царств живых организмов.
|
Царство |
Признаки |
Примечания |
|
Viroidobionta |
Неклеточное строение, инфекционность с помощью белка, активирующего участки генома хозяина; нуклеиновая кислота отсутствует (п/ц Pryonae) или представлена короткой РНК (п/ц Viroidae). |
Возможно, являются дегенерирующими потомками витоспор. В таком случае, первичная форма организации - вироидная, вторичная - прионная. |
|
Birnabionta |
Неклеточное строение; геном представлен двухцепочечной РНК; стратегия реализации: (2) РНК - (+) РНК - белок. |
|
|
Monornabionta |
Неклеточное строение; геном - одноцепочечная РНК; стратегия реализации: (-) РНК - (+) РНК - белок (п/ц Negarnaviridae), (+) РНК - (-) РНК - (+) РНК - белок (п/ц Posirnaviridae) или совмещение обеих стратегий (п/ц Arenoviridae). |
Информация на счет ареновирусов требует уточнения. |
|
Revertobionta |
Неклеточное строение; имеет место обратная транскрипция с помощью ревертазы; в вирионе присутствует РНК (п/ц Retroviridae) или ДНК (п/ц Hepadnaviridae). |
Монофилетичность практически очевидна (во всяком случае, возникли из схожих структур схожим способом), но стратегия реализации генома разная в разных подцарствах. |
|
Monodnabionta |
Неклеточное строение; геном представлен одноцепочечной ДНК. |
Вероятно, геном может быть как позитивным, так и негативным. |
|
Bidnabionta |
Неклеточное строение; геном представлен двухцепочечной ДНК, голой (п/ц Plasmidae) или покрытой капсидной оболочкой (п/ц Bidnaviridae); некоторые характеризуются автономностью репликации и транскрипции, отсутствием сплайсинга, большой величиной генома (п/ц Poxviridae). |
Возможно, поксвирусы происходят от дегенерировавших бактерий или от потомка прогенота с одним типом нуклеиновой кислоты и без рибосомного комплекса. |
|
Bacteriobionta |
Здесь и далее - клеточное строение (имеются рибосомные комплексы); прокариотический тип организации клетки; в составе клеточной стенки - муреин, РНК-полимераза из 4-8 субъединиц; строение коккоидное или сводимое к нему (п/ц Eubacteria), спиральное со сложной опорно-двигательной основой (п/ц Spirobacteria) или сложное мицелиального или близкого к нему типа (п/ц Actinobacteria). |
Возможно, подцарствам следует придать статус самостоятельных царств. |
|
Microsporidiobionta |
Эукариотический тип организации; рибосомы эубактериального типа, митохондрии и половой процесс отсутствуют. |
По происхождению, возможно, связаны с эубатериями, эукариотическую организацию приобрели независимо. |
|
Archaebionta |
Прокариотический тип организации клетки; эфиры глицерина и фитанола в составе липидов мембраны, кислые полисахариды, белки или псевдомуреин в стоставе клеточной стенки, РНК-полимераза из 9-12 субъединиц, многократно повторяющиеся нуклеотидные последовательности и интроны эукариотического типа в геноме. |
Вместе с настоящими эукариотами представляют собой отдельный ствол эволюции клеточных форм жизни, противопоставленный эубактериям. |
|
Archaezoobionta |
Эукариоты, митохондрии и половой процесс отсутствуют; может иметься бактериальный симбионт, выполняющий функцию митохондрий. |
Эта и дальнейшие группы животных относятся к настоящим эукариотам. |
|
Ciliobionta |
Кристы митохондрий истинно трубчатого строения; половой процесс - конъюгация; ундулиподии многочисленные (реснички). |
|
|
Pedinellobionta |
Хромосомы кольцевые (п/ц Dinoflagellata) или линейные; цитостом сзади; кристы ветвятся и анастомизируют (п/ц Myxobionta) или имеют вид пузырьков на тонких ножках; жгутикование часто гетероконтное, тогда жгутики обычно часто имеют трехразделенные трубчатые мастигонемы (п/ц Heterocontae), возможно также стефаноконтное жгутикование (п/ц Stephanocontae). |
Вероятно, кристы митохондрий близки к исходному типу. Между гетероконтами и миксобионтами, вероятно, есть промежуточная группа - ваушериевые. Принадлежность стефаноконт к данному царству точно не выявлена . |
|
Sporobionta |
Обычно имеются коноид, роптрии, полярное кольцо; тип крист близок к трубчатому; жгутикование каудоконтное (п/ц Caudocontae) или изоконтное (п/ц Coccidiomorpha). |
Родственные связи с другими группами трубчатокристовых и между собой не ясны. Возможно, сборная группа, которую следует либо разделить на два царства, либо раскидать по уже имеющимся. |
|
Cnidosporidiobionta |
Трубчатые кристы; многоклеточные споры со стрекательными капсулами. |
Родственные связи с другими группами трубчатокристовых не ясны. |
|
Discocristobionta |
Закрытый ортомитоз с сохранением ядрышка; полового процесса нет; цитостом впереди; кристы дисковидные. |
Группа, возможно, родственна пластинчатокристовым. |
|
Cryptobionta |
Кристы гребневидные; цитостом впереди; ортомитоз с разрушением ядрышка; в качестве пластид - эукариотический эндосимбионт; бичевидный и лентовидный жгутики. |
Родственные связи с другими пластинчатокристовыми не ясны; по жгутикованию, возможно, близки к исходному типу пластинчатокристовых. |
|
Phytobionta |
Кристы гребневидные; пластиды желтые (п/ц Haptophyta) или зеленые; клеточное деление без фрагмопласта и клеточной пластинки (п/ц Phycophyta) или с образованием оных (п/ц Metaphyta); жгутикование у высших растений изоконтное, мультиконтное или отсутствует, причем утрата жгутиков полифилетична. |
В эволюции растений четко прослеживается прогрессивная эволюция. Принадлежность гаптофитов к растениям сомнительна. Зеленые водоросли, возможно, распадаются на несколько подцарств. Конъюгаты близки к высшим растениям. |
|
Rhodobionta |
Кристы гребневидные; жгутиковая стадия обычно отсутствует; пластиды обычно красные (п/ц Rhodophyta); характеристика второго подцарства (п/ц Glaucophyta) нам не известна. |
Родственные связи с другими пластинчатокристовыми не ясны; возможно, родственны грибам. |
|
Mycobionta |
Кристы гребневидные; в клеточных стенках присутствует хитин; жгутик задний (п/ц Promycota) или отсутствует (п/ц Eumycota). |
Родственны животным. |
|
Zoobionta |
Кристы гребневидные; жгутиковая стадия обычно заднежгутиковая; клеточных стенок нет; одноклеточные или колониальные формы (п/ц Protozoa) либо многоклеточные организмы, имеющие эмбриогенез с бластулой (п/ц Metazoa). |
Как и в царстве растений, прослеживается прогрессивная эволюция. |
ПРИЛОЖЕНИЕ 2. Перечень высших таксонов современных живых организмов.
Ц. Viroidobionta
п/ц. Viroidae
п/ц. Pryonae
Ц. Birnabionta
Ц. Monornabionta
п/ц. Negarnaviridae
п/ц. Arenoviridae?
п/ц. Posirnaviridae
Ц. Revertobionta
п/ц. Retroviridae
п/ц. Hepadnaviridae?
Ц. Monodnabionta
Ц. Bidnabionta?
п/ц. Plasmidae
п/ц. Bidnaviridae
п/ц. Poxviridae
Ц. Bacteriobionta?
п/ц. Eubacteria
т. Eubacteria
к. Clostridiobacteria
к. Thiopneutes
к. Azotobacteria
к. Pseudomonadae
к. Aёroendospora
к. Aphragmobacteria
к. Micrococci
к. Omnibacteria
к. Rikketsiomorpha
т. Oxybacteria
к. Hemobacteria
к. Tetrapirrobacteria
к. Proplastidae
п/ц. Spirobacteria
т. Spirochaetae
т. Spiroplasmae
п/ц. Actinobacteria
т. Myxobacteria
т. Actinobacteria
Ц. Microsporidiobionta
Ц
. Archaebiontaт. Methanobacteria
т. Halobacteria
т. Acidobacteria
Ц. Archaezoobionta
т. Caryoblastea
т. Diplomonada
т. Trichomonada
Ц. Ciliobionta
т. Kinetofragminophora
к. Gymnostomata
к. Vestibulifera
к. Hypostomata
к. Suctoria
т. Oligohymenophora
к. Hy
к. Peritricha
т. Polyhymenophora
Ц. Pedinellobionta
п/ц. Dinoflagellata
п/ц. Heterocontae
т. Actinophryidae
+ Ciliophryidae
т. Oomycota
к. Hyphochytridiomycetes
к. Traustochytridiomycetes
к. Oomycetes
к. Labyrinthulomycetes
т. Chromista?
к
к. Prasinophyceae
к. Raphidophyceae
к. Xantophyceae
к. Chrysophyceae
к. Phaeophyceae
к. Bacillariophyceae?
п/ц. Myxobionta
т. Rhyzopoda?
к. Lobosida
к. Acanthamoebida
к. Radiolaria
к. Euheliozoa
+ Centrohelidia
т. Haemosporidida?
к. Haemosp
к. Haplosporidia?
т. Myxomycota
к. Foraminifera?
к. Protostellida
к. Discostellida
к. Myxogastrea
к. Plasmodiophorida?
т. Mastigophora?
к. Chlororarchniophyceae
к. Rhyzomastigida?
к. Eumastigida
т. Slopalinida
к. Opalinida
к. Pseudosporea
п/ц
. Stephanocontae?т. Oedogoniophyta
т. Siphonophyta
Ц. Sporobionta?
п/ц. Coccidiomorpha
т. Coccidiida
п/ц. Caudocontae
т. Gregarinida
к. Spiromonadida
к. Eugregarina
Ц. Cnidosporidiobionta
т. Myxosporea
т. Actinosporea
Ц. Discocristobionta
т. Euglenophyta
т. Kinetoplastida
т. Pseudociliata
т. Heterolobosea
к. Acrasiomycetes
к. Schizopirenidae
к. Nucleariidae?
Ц. Cryptobionta
Ц. Phytobionta
п/ц. Haptophyta?
т. Haptophyceae
п/ц. Phycophyta?
т. Micromonadophyceae
т. Pleurastophyceae
т. Ulvophyceae
т
п/ц. Metaphyta
т. Zygnematophyta?
к. Desmidiopsida
к. Mesotaeniopsida
к. Gonadozygopsida
к. Zygnematopsida
т. Klebsormidiophyta
+ Chaetophorales?
т. Anthoceratophyta
т. Charophyta
т. Hepatophyta
к. Marchantiopsida
к. Jungermanniopsida
т. Br
yophytaк. Sphagnopsida
к. Andreopsida
к. Bryopsida
т. Lycopodiophyta
к. Lycopodiopsida
к. Selaginellopsida
т. Tmesophyta
т. Isoёtophyta
т. Equisetophyta
т. Polypodiophyta?
к. Ophioglossopsida
к. Marattiopsida
к. Polypodiopsida
к. Marsileopsida
к
т. Cycadophyta
т. Ginkgophyta
т. Pinophyta
т. Tracheophyta
к. Ephedropsida
к. Welwitschiopsida
к. Gnetopsida
к. Magnoliidae
+ Ranunculidae
к. Hamamelidopsida
к. Caryophyllopsida
к. Dilleniopsida
к. Rosopsida
к. Asteropsida
к. Liliopsida
+ Nympheaceae
Ц. Rhodobionta
т. Glaucophyta?
т. Rhodophyta
к. Bangiophyceae
к. Florideophyceae
Ц. Mycobionta
п/ц. Promycota
т. Chytridiomycota
п/ц. Eumycota
т. Zygomycota
т. Ascomycota
к. Hemiascomycetes
к. Plectomycetes
к. Pirenomycetes
к. Discomycetes
к.
т. Basidiomycota
к. Exobasidiomycetes
к. Hymenomycetes
к. Gasteromycetes
к. Teliosporomycetes
к. Heterobasidiomycetes
Ц. Zoobionta
п/ц. Protozoa
т. Choanoflagellata
п/ц. Metazoa
т. Placozoa
т. Plathelmintes
к. Acoelida
к. Xenoturbellida
к. Udonellida
к. Aspidogastrida
к. Euturbellida
к. Monogenea
к. Cestoda
к. Cestodaria
к. Trematoda
к. Mesozoa?
т. Nemathelmintes
к. Gastrotricha?
к. Nematoda
т. Nemertini
т. Spongia
к. Parenchimulozoa
к. Amphiblastulozoa
т. Coelenterata
к. Hydrozoa
к. Scyp
к. Anthozoa
т. Kamptozoa
т. Dinofilida
т. Echiurida
т. Myzostomida
т. Sipunculida
т. Annelida?
к. Polychaeta?
к. Olygochaeta
к. Hirudinea
т. Mollusca
к. Monoplacophora
к. Solenogastres
к. Caudofovea
к. Loricata
к. Lamellibranchia
к. Gastropoda
к
к. Cephalopoda
т. Tardigradomorphes
к. Tardigrada
к. Pentastomida
к. Onychophora
т. Arthropoda?
к. Crustacea
к. Chelicerata
к. Pantopoda
к. Diplopoda
к. Chilopoda
к. Pauropoda
к. Symphyla
+ Collembola
к. Diplura
к. Protura
к. Insecta
т
. Podaxoniaк. Phoronoidea
к. Bryozoa
т. Cephalorhyncha
к. Rotatoria?
к. Acanthocephala?
к. Kinorhyncha
к. Loricifera
к. Priapulida
к. Nematomorpha
т. Ctenophora
т. Brachiopoda
т. Chaetognatha
т. Pogonophora?
т. Echinodermata
к. Crinoidea
к. Holothurioidea
к. Echinoidea
к. Asteroidea
к. Concentricycloidea
к. Ophiuroidea
т. Hemichordata
к. Pterobranchia
к. Enteropneusta
т. Chordata
к. Tunicata?
к. Acrania
к. Agnatha?
к. Teleostei
+ Holosteimorpha
к. Holocephali
к. Elasmobranchii
+ Acipenseriformes
к
. Sarcopterigii+ Dipnoi? + Polyptera?
к. Amphibia?
к. Sinapsida
к. Anapsida
к. Diapsida
Адрианов А., Малахов В. Сравнительно-морфологический анализ организации головохоботных червей, филогения и система типа Cephalorhyncha // Зоологический журнал. - т. 74, вып. 4-8.
Адрианов Р. Филогенетическое положение мохообразных и родственных им групп в контексте происхождения и эволюции высших растений: Автореферат доклада на конференции студентов, аспирантов и преподавателей КГУ. - Калининград: 1996. - 5 стр.
Айала Ф., Кайгер Дж. Современная генетика - в 3-х т. - М.: Мир, 1987.
Айкхорн С., Рейвн П., Эверт Р. Современная ботаника - в 2-х т. - М.: Мир, 1990.
Антонов А., Боброва В. Молекулярно-биологические исследования происхождения покрытосеменных растений // Журнал общей биологии. - т. 56, №6.
Бардунов Л. Древнейшие на суше. - Новосибирск: Наука, 1984.
Беклемишев К. Зоология беспозвоночных. - М.: МГУ, 1979.
Берг Л. Труды по теории эволюции. - Л.: Наука, 1977.
Богомолов С. Сокращение раннего эмбриогенеза и дивергентная эволюция у некоторых примитивных многоклеточных животных // Сб. “Темп индивидуального развития животных и его изменение в ходе эволюции”. - М.: 1968.
Вагин В. Телобластическое развитие у артикулят как гетерохрония и его связь с эволюцией спирального дробления // Сб. “Темп индивидуального развития животных и его изменение в ходе эволюции”. - М.: 1968.
Васильев И. Живое из неживого. - М.: Знание, 1965.
Вернадский В. Биосфера. - М.: Наука, 1967.
Вирусология, в 3-х т. / ред. Филдс Б., Найн Д. - М.: Мир, 1989.
Водоросли (справочник). - Киев: Наукова думка, 1989.
Войнович Г. Возникновение и развитие жизни на Земле. - М.: Наука, 1988.
Все живое на Земле произошло из нефти! // Комсомольская правда. - 1999. - 11 марта.
Гарибова Л., Горбунова Н., Сидорова И., Сизова И. Низшие растения. - М.: МГУ, 1975.
Гвоздев В. Подвижная ДНК эукариот. // Соросовский образовательный журнал. - 1998, №8. - Стр. 8 - 21.
Генетика. - М.: 1991.
Георгиевский А. Проблема преадаптации. - Л.: Наука, 1974.
Глазер В. Гомологическая генетическая рекомбинация. // Соросовский образовательный журнал. - 1998, №7. - Стр. 13 - 21.
Глазер В. Генетическая рекомбинация без гомологии: процессы, ведущие к перестройке в геноме. // Соросовский образовательный журнал. - 1998, №7. - Стр. 22 - 29.
Глобальный эволюционизм / ред. Фасенковой и др. - М.: Ифран, 1994.
Грант В. Эволюционный процесс. - М.: Мир,1991.
Грант В. Эволюция организмов. - М.: Мир, 1980.
Догель В. Зоология беспозвоночных. - М.: 80.
Дарвин Ч. Происхождение видов. - М.: ГИСХЛ, 1952.
Жданов В. Эволюция вирусов. - М.: Медицина, 1966.
Жизнь животных - в 7-и т. - М.: Просвещение.
Жизнь - как она возникла? - New York: WBATS, 1985.
Жизнь растений - в 6-и т. - М.: Просвещение.
Жуковский П. Ботаника. - М.: ВШ, 1964.
Захваткин А. Сравнительная эмбриология низших беспозвоночных. - М.: Современная наука, 1949.
Зусман И. Вопросы эволюции эмбриогенеза животных. - М.: Знание, 1973.
Иванов А. Происхождение многоклеточных животных. - Л.: Наука, 1968.
Кейлоу П. Принципы эволюции. - М.: Мир, 1986.
Красилов В. Нерешенные проблемы теории эволюции. - Владивосток: 1986.
Красовский В., Шкловский И. Возможное влияние вспышек сверхновых на эволюцию жизни на Земле. // ДАН СССР. - 1957, т. 116, №2. - Стр. 197-199.
Кулаев И. Происхождение эукариот. // Соросовский образовательный журнал. - 1998, №5. - Стр. 17 - 22.
Курс низших растений. - М.: ВШ, 1981.
Лемеза Н., Шуханов Н. Малый практикум по низшим растениям. - Минск: Унiверсiтэцкае, 1994.
Лима де Фариа А. Эволюция без отбора. - М.: Мир, 1991.
Малахов В., Реунов А. Эволюция строения сперматозоидов у беспозвоночных // Успехи современной биологии. - т. 113, вып. 1. - Стр. 3.
Маргелис Л. Роль симбиоза в эволюции клетки. - М.: Мир, 1983.
Медведев Н. Беседы по биологии пола. - Минск: 1976.
Мирабдуллаев И. О происхождении и положении грибов в системе органического мира // Успехи современной биологии. - т. 114, вып. 1.
Мирабдуллаев И. Проблема происхождения эукариот // Успехи современной биологии. - т. 107, вып. 3.
Мирабдуллаев И. Проблемы классификации живого на уровне царств // Журнал общей биологии. - т. 50, №6.
Мирабдуллаев И. Рибосомы, кристы и филогения низших эукариот // Издание АН СССР, серия Биология. - 1989. - №5.
Молчан И. Космизация курса общей биологии в системе опережающего образования и науки: прогнозы Живой Этики. // Три ключа (педагогический вестник). - 1998, вып. 2. - Стр. 36 - 51.
Московский А. Существует ли научная альтернатива дарвинской концепции эволюции? // Три ключа (педагогический вестник). - 1999, вып. 3. - Стр. 49 - 54.
Назаров В. Учение о макроэволюции. - М.: Наука, 1991.
Наумов Н., Карташев Н. Зоология позвоночных - в 2-х т. - М.: ВШ, 1979.
Николаев Г. Магнитное поле Земли слабеет. Опасны ли последствия этого? // Наука и жизнь. - 1991, №1. - Стр. 44-50.
Опарин А. Возникновение жизни на Земле. - М.: 57.
Опарин А. Возникновение и начальное развитие жизни. - М.: Медицина, 1966.
Павлинов И. Кладистический анализ. - М.: МГУ, 1990.
Пахомова М. ДНК водорослей. Строение ДНК и положение организмов в системе. - М.: МГУ, 1972.
Петров Ю. Основные этапы эволюции циклов развития у водорослей // Сб. “Проблемы филогении низших растений”. - М.: Наука, 1974.
Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция. - М.: Мир, 1986.
Самовар А. Происхождение полового процесса и эукариот - взаимосвязанные проблемы? // Журнал эволюционной биохимии и физиологии. - №28. - С. 91.
Северцев А. Направленность эволюции. - М.: МГУ, 1990.
Седова Т. Типы митоза и закономерности их распределения в различных отделах водорослей // Ботанический журнал. - т. 80.
Симпсон Дж. Темпы и формы эволюции. - М.: ИИЛ, 1948.
Соболев Д. Начала исторической биогенетики. - Симферополь: Госиздат Украины, 1924.
Стеббинс Дж., Айала Ф. Эволюция дарвинизма // В мире науки. - №8, 1984. - С. 26-35.
Тихоненко Т. Роль вирусов в обмене генетической информацией. - М.: Знание, 1980.
Хесин Р. Непостоянство генома. - М.: Наука, 1984.
Хржановский В., Пономаренко С. Ботаника. - М.: Агропромиздат, 1988.
Черникова И. Глобальный эволюционизм. - Томск: ТУ, 1987.
Четвериков С. О некоторых моментах эволюционного процесса с точки зрения современной генетики (1926) // Классики современной генетики. - Л.: Наука, 68. - С. 133-170.
Шкловский И. Вселенная, жизнь, разум. - М.: Наука, 1973.
Шляков Р. Печеночные мхи: морфология, филогения, классификация. - Л.: Наука, 1975.
Шпет Г. Темп роста в онтогенезе представителей древних групп животных // Сб. “Эволюция темпов эволюционного развития животных”. - М.: 1977.
Эволюция генома. - М.: 1986.